
|
Wednesday, June 14, 2006
Uhh.. No time to sleep cos if ya sleep ya don't eat... Gotta hold heat just to make ends meet.. - T3
I got to thinking about flies again, partially because Ron Davis came and gave a talk at our department and partially because I saw a couple papers in Nature showing sleep regulation is mediated by protein kinase A (PKA) signaling in the mushroom bodies. You may be surprised to find out that flies sleep, but they at least do something like sleeping where they sit very still and it's harder to arouse them. Then once I started digging in I found yet another paper in Current Biology that came out within two days of the Nature papers implicating serotonin 5-HT1A receptors in the mushroom bodies. 5-HT1A receptors and PKA are linked through the cyclic AMP (cAMP) signaling pathway. PKA is also referred to as cAMP-dependent protein kinase. Where else have I seen all these cats before? Ah yes. Here they are inititating memory formation in Kandel's Nobel lecture about the mechanisms of long-term facilitation in another invertebrate (the sea slug). 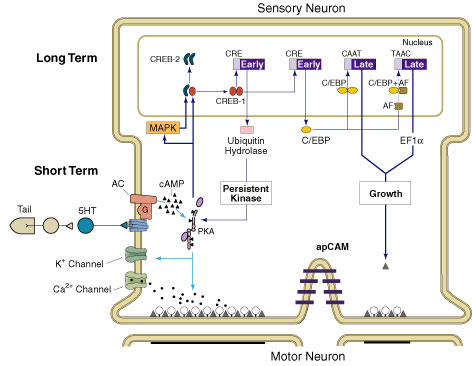 In fact, the PKA signaling pathway in the mushroom bodies is known to be of major importance for memory formation in fruit flies as well. Two of the earliest discovered fly memory mutants, rutabaga and dunce, ended up lacking PKA pathway proteins, and at least the rutabaga memory phenotype can be rescued by inducing the missing protein just in adult mushroom bodies. I keep talking about mushroom bodies. Mushroom bodies are a bilateral tripartite structures found in the insect brain. Most of the mushroom bodies are made up of the axons of Kenyon cells. The Kenyon cells are all collected together in a big round hunk of tissue positioned to the dorsal-posterior. A tract called the peduncle shoots down and forward and a little lateral and then abruptly splits into lobes. For simplicity's sake I will pretend that there are only alpha, beta, and gamma lobes. The alpha lobe shoots straight up from the branching point producing a vertical lobe. The horizontal offshoot from the branching point goes medial and consists of beta and gamma lobes which skilled fly neuroanatomists can tell apart using arcane fly researcher magic. Actually I think the gamma lobe is just like in front of the beta lobe or something. Mushroom bodies are known to be important for memory (especially long-term memory) because of things like the aforementioned rutabaga rescue experiments and because you can screw up memory by cooling them or ablating them with hydroxyurea. Also, they are kind of in a good spot to encode the type of memory most people study in fruit flies (aversive olfactory conditioning). They are two or three synapses from the olfactory receptors and they receive neuromodulatory input from transmitters proposed to be involved in shock representation. All of these papers use the Gal4-UAS system, so I ought to lay it out. If fly researchers want to affect a gene's expression in a cell-type specific fashion they can just pull a fly Gal4 line off the shelf. Gal4 is a transcription factor that has been inserted in multiple spots in the fly genome producing flies that make Gal4, say, just in certain Kenyon cells or in the antennal lobe, or in the protocerebrum, or wherever. This happens because the Gal4 sequence is being driven by some other gene's regulatory region (promoter). A lot of times, they don't even know which gene's promoter is being co-opted, but hey it works. Gal4 has the property of activating transcription of genes that are controlled by an Upstream Activating Sequence (UAS). So you have to make a fly that has whatever gene you're interested in controlled by a UAS. Then you grab a Gal4 strain that has expression in the cell-type you're interested in and cross'em. Gal4 + UAS = your gene expressed solely in the tissue of interest. Pitman et al. started out by inserting a temperature-sensitive inhibitor of neurotransmitter release (shibire) after a UAS and trying it out in different Gal4 lines to find sleep centers. They found a few lines with reduced sleep (short-sleep lines, SSLs), and then looked to see where Gal4 was being expressed by using the same Gal4 lines but crossing them with UAS-GFP strains instead of the neural silencer. They mostly found expression in the mushroom bodies, so they tried out a couple more known mushroom body lines (c309 and 30Y), and these showed short-sleep phenotypes too. The overall idea here is that mushroom body activity is sleep-promoting. Interestingly, a c309-PKA line also reduced sleep. So turning off neurotransmitter release seems to have the same effect as turning up PKA activity. Also, sleep-reducing changes were associated with a reduction in lifespan. I will use this finding to cynically sensationalize any further findings regarding sleep regulation by making them seem a matter of life or death. Since the sleep regulation machinery overlaps so much with synaptic plasticity and memory pathways, I will use this as an excuse to say things like "Learning can kill you!" or "Learning can save your life!" depending on which way I interpret the rest of this data. Joiner et al. did a similar set of experiments, but instead of turning off neurotransmitter release in certain Gal4 lines, they expressed a constitutively active version of PKA. They found sleep phenotypes in two lines: 201Y and c309 again. It's rare to see an experimental result independently replicated period, let alone on the next page of the same journal, but here we have it. c309-PKA flies showed reduced sleep in Joiner et al.'s hands too. 201Y-PKA flies showed an increased sleep phenotype. The authors of this paper were a little more subtle with their histology and determined that c309 and 201Y did indeed both have mushroom body expression patterns, but they have a complementary pattern within the alpha and beta lobes. 201Y is found in gamma lobes and the core region of the alpha and beta lobes while c309 is found in the gamma lobes and not in the core region of the alpha and beta lobes. These folks also have a mushroom body Gal4 line with similar expression pattern to c309 that can be turned on in response to a drug (RU486), so they could isolate the adult contribution rather than the developmental contribution of PKA activity. This manipulation was also associated with reduced sleep. Tissue and time-specifically expressing potassium channels to inhibit mushroom body activity or sodium channels to activate led to increased and decreased sleep respectively. But didn't we just learn that mushroom body activity is sleep-promoting? In fact, this paper also contains the finding that mushroom bodies leads to less sleep, but they note that other manipulations can produce much larger changes. Mushroom bodies probably contain populations that do both and when you ablate them you are canceling out a little bit of both. I got interested in this idea and started looking for indicators of what might be different between the two populations. The major distinction seems to be that PKA activity (and thus more activity?) in the core of the alpha and beta lobes is sleep promoting while PKA outside of the core is sleep reducing. I found one paper that shows a GABA receptor subunit is primarily found in the core of alpha and beta lobes. GABA is normally an inhibitory input, so having GABA receptors throws a minus sign in the equation, but these are receptors not neurotransmitter, so this only explains a differential response to external stimuli between core and non-core. Another paper shows that calcium/calmodulin-dependent kinase II (CaMKII, an important synaptic plasticity molecule) is mostly non-core, as are two important PKA signaling molecules, Leonardo and Rutabaga. This indicates to me that it is a little unnatural to be expressing active PKA in the core (and thus promoting sleep). This leads me to believe that endogenous PKA activity is probably sleep reducing. Finally, there's this Yuan et al. paper from the same lab as the Joiner et al. paper. In this paper the focus is on serotonin receptor subtypes. The authors produced disruptions in each of the drosophila 5HT receptor subtypes and looked for sleep phenotypes. Only the 5HT-1A receptor disruption had an effect on sleep, producing a short sleep phenotype. Reinstating 5HT-1A using a pan-neuronal Gal4 driver and UAS-5HT-1AR, rescued sleep. Also, expressing the receptor using the Gal4 MB-Switch driver (the one that can be turned on with RU486) just in adult mushroom bodies rescued sleep. This pretty much ruins my core versus non-core theorizing because the aforementioned "pan-neuronal" driver has more intense expression in the core while the MB-Switch driver is preferentially non-core, but the outcome is the same for expressing 5HT-1A receptors in both. When I read all of these at once I had hoped to come up with some grand overarching theory of memory, sleep, and cyclic AMP signaling, but, alas, the data isn't that neatly sewn up. It seems like serotonin and PKA are having opposite effects. The story would fit more neatly into my scheme if PKA activity in the mushroom bodies was sleep promoting, and it is in certain neuronal populations. If I combine this with Pitman et al.'s general finding that mushroom body activity seems to be sleep-promoting and ignore the inconvenient data points from Joiner et al., then I could say something like "learning induces signal cascades that produce greater mushroom body activity and promote sleep." Then I could tie it in with theories about the role of network activity (like sharp-wave ripples) in the mammalian brain during sleep. Turns out it isn't that simple. Any ideas? How do you like reading a blog post without the answer at the end? |
  Razib's Home Page GNXP Archives Interviews Blogroll Principles of Population Genetics Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism
Hello
Movable Type archives August 11,2002 August 18,2002 August 25,2002 September 01,2002 September 15,2002 October 20,2002 December 08,2002 December 22,2002 December 29,2002 January 05,2003 January 12,2003 January 19,2003 January 26,2003 February 02,2003 February 09,2003 February 16,2003 February 23,2003 March 02,2003 March 09,2003 March 16,2003 March 23,2003 March 30,2003 April 06,2003 April 13,2003 April 20,2003 April 27,2003 May 04,2003 May 11,2003 May 18,2003 May 25,2003 June 01,2003 June 08,2003 June 15,2003 June 22,2003 June 29,2003 July 06,2003 July 13,2003 July 20,2003 July 27,2003 August 03,2003 August 10,2003 August 17,2003 August 24,2003 August 31,2003 September 07,2003 September 14,2003 September 21,2003 September 28,2003 October 05,2003 October 12,2003 October 19,2003 October 26,2003 November 02,2003 November 09,2003 November 16,2003 November 23,2003 November 30,2003 December 07,2003 December 14,2003 December 21,2003 December 28,2003 January 04,2004 January 11,2004 January 18,2004 January 25,2004 February 01,2004 February 08,2004 February 15,2004 February 22,2004 February 29,2004 March 07,2004 March 14,2004 March 21,2004 March 28,2004 April 04,2004 April 11,2004 April 18,2004 April 25,2004 May 02,2004 May 09,2004 May 16,2004 May 23,2004 May 30,2004 June 06,2004 June 13,2004 June 20,2004 June 27,2004 July 04,2004 July 11,2004 July 18,2004 July 25,2004 August 01,2004 August 08,2004 August 15,2004 August 22,2004 August 29,2004 September 05,2004 September 12,2004 September 19,2004 September 26,2004 October 03,2004 October 10,2004 October 17,2004 October 24,2004 October 31,2004 November 07,2004 November 14,2004 November 21,2004 November 28,2004 December 05,2004 December 12,2004 December 19,2004 December 26,2004 January 02,2005 January 09,2005 January 16,2005 January 23,2005 January 30,2005 February 06,2005 February 13,2005 February 20,2005 February 27,2005 March 06,2005 March 13,2005 March 20,2005 March 27,2005 April 03,2005 April 10,2005 April 17,2005 April 24,2005 May 01,2005 May 08,2005 May 15,2005 May 22,2005 May 29,2005 June 05,2005 June 12,2005 June 19,2005 June 26,2005 July 03,2005 July 17,2005 August 07,2005 Blogspot archives June 2002 July 2002 August 2002 September 2002 October 2002 November 2002 December 2002
10 questions for....
Parag Khanna James Flynn Jon Entine Gregory Clark György Buzsáki Heather Mac Donald Bruce Lahn A.W.F. Edwards Luigi Luca Cavalli-Sforza Joseph LeDoux Matthew Stewart Charles Murray James F. Crow Adam K. Webb Justin L. Barrett David Haig Judith Rich Harris Ken Miller Dan Sperber Warren Treadgold Armand M. Leroi John Derbyshire
Blogs
The GiveWell Blog Your Religion Is False Colby Cosh Steve Hsu Audacious Epigone Catallaxy Files Inductivist 2 Blowhards Genetic Future Agnostic Steve Sailer Dienekes Derek Lowe Razib Khan Razib at Comment is Free Secular Right Glenn Reynolds Jim Miller Kevin McGrew John Hawks Peter Fost Randall Parker Less Wrong Charles Murray Carl Zimmer EconLog Marginal Revolution
Principles of Population Genetics
Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism   Policies Terms of use © http://www.gnxp.com Razib's total feed: |