|
Leave print-view
Sunday, August 06, 2006
Grind ya teeth and just roll with it. Don't risk it. Fuck around and be a statistic. - Ghostface genius re-post: This article by McCoy and Platt in the September 2005 issue of Nature Neuroscience is an extension of past work and a clarification of what was previously observed. In the past study, they found reward-sensitive neurons in the posterior cingulate cortex. In this more recent study, the neurons are utility-sensitive. The difference is between measuring the more objectively available idea of reward size and the more subjective idea of utility. To quote the commentary by Daeyeol Lee, "In economics, the numerical measure of an individual's preference of subjective value for an object is referred to as utility."
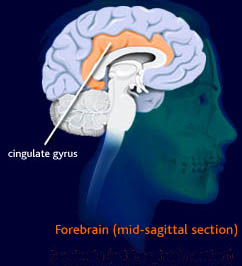
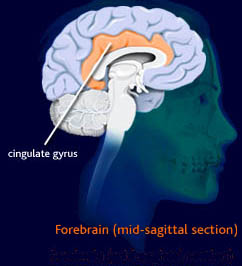
Let's back up a couple steps, one step for anatomy and one for the previous results. First, let's get an image of where this posterior cingulate (CGp) is. Many are familiar with the corpus callosum, the giant bundle of white matter that connects the hemispheres. The cingulate basically wraps around this structure over the top. So if you were to take a brain and cut it in half lengthwise and pretend it was a rainbow, the third ventricle/thalamus/septum pellucidum and whatnot would be red-orange, the corpus callosum would be yellow, the cingulate would be green, the rest of the cortex would be blue, and your psychic aura would be indigo and violet. The portion of the cingulate that's further back toward the brain stem is the CGp. I'm not sure, but I'm betting there is a lot of vagueness regarding the boundary between the anterior and posterior. The illustration below is not color-coded in the way I described. I got it here and it from there it points to here:
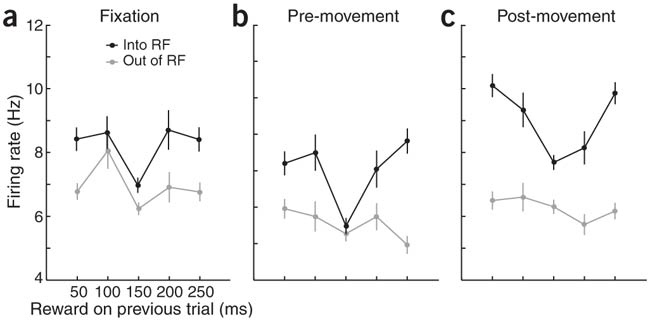
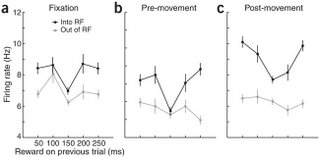
Now waaaay back in 2003 is when McCoy et al. recorded reward signals in the CGp. They used single-neuron recording in a saccade (eye-movement) task to measure this. In order to get some delicious juice the monkeys had to stare at an asterisk in the middle of a screen (the fixation point), wait for a target spot elsewhere on the screen to light up, wait for the fixation point to disappear, then move their eyes to the target spot at which point they received said delicious juice. Every 50 trials they would alter the amount of juice received for a movement to the target spot. The saccade behavior indicated that the monkeys were noticing the difference. On trials when the juice reward was small, they would shift their eyes very quickly so as to have the most time to lap up as much juice as possible.
In addition to behavior reflecting changes in reward size, neurons in the CGp also seemed to be getting the picture. Two brands of neurons changed their firing rate in accordance with changes in reward size. One set signaled a short time after the actual eye-movement, and the other set signaled a short time after the reward was administered. A given neuron could increase or decrease firing at one of these times. Increasing the reward size seemed to be increasing the gain on the signal such that cells that usually increased firing would increase further with larger rewards, but ones that suppressed firing would decrease further with larger rewards. The authors interpreted the first firing period (after the movement) as predictive and the second as representing the actual value of the reward.
The monkey's saccade is thus analogous to going to bed on the night before Xmas and the firing that follows is the equivalent of dreams of sugarplums dancing in their heads. The firing after reward is reflects the actual receipt of sugarplums. If the neuron was of the increasing variety, then firing might be really high for one of those gigantic Pixie Sticks and quite lower for a protein bar. There is also an entirely statistically separate signal corresponding to no reward at all when one is expected. This is the "stop believing in Santa Claus" signal, and a neuron that usually decreased firing with lower rewards could still start firing like crazy when the reward is non-existent which goes to show that the signal is a special one. I don't know how this thing got so Xmas-themed so quickly. I must not have invoked the spirit of Ghostface hard enough.
So here's the deal with the newer paper. Maybe there are some kids that really love protein bars more than pure sugar. The reward scale would be all screwed up if we simply tried to measure it objectively. From the last set of experiments we can't tell whether the neurons are signaling something like "juice received" or something more subjective like "how awesome the next trial is gonna be cos I love juice." This is where the gambling comes in. McCoy and Platt cleverly found a way to rule out the "juice received" signal theory.
The first thing they did was to show that macaques in this task are exceptionally risk-prone. They looove to gamble. These monkeys are not wearing their safety belt while driving to Staten Island to try to peddle weight in Theodore Unit's territory. This was discovered using the same saccade task with the addition of another choice for the target point. After fixation, two choices were available: 1) a certain target that always delivered the same amount of juice or 2) a risky target that sometimes delivered less and sometimes delivered more but averaged out to the same amount as the certain target. The monkeys overwhelmingly prefer the risky target. In fact, they prefer the risky target even if it delivers a little less juice overall than the certain target.
So you can see where this is going. What are neurons in the CGp doing? Staying exactly the same all the time since the same average reward is received or changing with the increased preference for the risky target? The latter is the case and the authors take this as evidence that these neurons are actually signaling expected utility. The "expected" is in there because this study is a lot more about those neurons that fire right after the saccade but before the reward. The effect of increased risk is to once again increase gain in this population, and also to increase spatial selectivity. These neurons have a particular direction of movement they are more happy firing about. Increased risk causes them to be even pickier about all of that.
The weird thing is that neurons actually increase their firing rate for the next trial following both of the non-average types of trials. After either an exceptionally low or high reward, that monkey really wants to go for the risky target again. I don't understand it. Seems sort of like buying a scratch-off ticket and losing, getting all determined to win your dollar back, and spending five more bucks on scratch-off tickets.
So we get a peek at something that seems subjective by measuring firing rates. That's pretty cool because the realm of the subjective is often a tough one to quantify. For those who wonder why their tax-payer dollars are being spent on figuring out that monkeys are big gamblers, here's the last word from the authors: "Neurophysiological studies of risk preferences, as reported here, may serve as an important model for probing neural processes that underlie pathological risk taking in individuals with addictions to drugs, sex, food or gambling."
References:
McCoy AN, Crowley JC, Haghighian G, Dean HL, Platt ML. (2003) Saccade reward signals in posterior cingulate cortex. Neuron 40:1031-1040.
Lee D. (2005) Neuroeconomics: making risky choices in the brain. Nat. Neurosci. 8:1129-1130.
McCoy AN, Platt ML. (2005) Risk-sensitive neurons in macaque posterior cingulate cortex. Nat. Neurosci. 8:1220-1227.
|