
|
Wednesday, May 31, 2006
This report is from today's London Independent. The source is the latest issue of Nature, so there will doubtless be other reports.
Update from Razib: Carl Zimmer and John Hawks have much more. One point that is really confusing about the Hobbits is their small brain size. Consider the graph to the left.  Human (hominids) brains keep getting larger, come rain or shine. The Hobbits, if they truly are what is claimed, radically reversed this trend. Convential explanations of island ecology seem a bit weak when set against this almost inevitable encephalization that seems to have characterized our lineage. Human (hominids) brains keep getting larger, come rain or shine. The Hobbits, if they truly are what is claimed, radically reversed this trend. Convential explanations of island ecology seem a bit weak when set against this almost inevitable encephalization that seems to have characterized our lineage.
I was poking around looking for something else and found this ESI special topics website. It updates every other month and provides interviews with the authors of the most highly cited papers at the time. I really think the "fronts" are interesting and could be useful. They give you a map of journal articles that seem to be forming a cluster and generating excitement within a certain field. Here's one for hippocampal GABA(A) receptors.
I'm hoping they do a map for the RNA decapping and degradation since they have identified it as a Fast Moving Front. They have an interview with Roy Parker about P-bodies and RNA degradation here. I've done a little bit on the topic before. The hot topic in the social sciences is apparently "Democracy". So check it out. ESI special topics: from Decapping to Democracy.
Given that some of the readers here are actual, like, scientists, this article on the relationship between marriage and scientific success may be of interest. Actually, the only interesting part is the intro:
Several years ago, Satoshi Kanazawa, then a psychologist at the University of Canterbury in Christchurch, New Zealand, analyzed a biographical database of 280 great scientists--mathematicians, physicists, chemists, and biologists. When he calculated the age of each scientist at the peak of his career--the sample was predominantly male--Kanazawa noted an interesting trend. After a crest during the third decade of life, scientific productivity--as evidenced by major discoveries and publications--fell off dramatically with age. When he looked at the marital history of the sample, he found that the decline in productivity was less severe among men who had never been married. As a group, unmarried scientists continued to achieve well into their late 50s, and their rates of decline were slower. The rest is a bunch of quotes from dudes bitching about how they can't meet women; read at your own risk. Related: Kanazawa on domestic violence and the sex ratio
Tuesday, May 30, 2006
Jeez, how long has it been since we had a good chat about comely girls? Via Dienekes, this article shows that in a sample of early 20-something girls at a Scottish university, daughters of happily married parents (HM) had more attractive and feminine faces than those whose parents either had separated pre-puberty (S) or had remained married but unhappily (UM). Color pictures in PDF here (L-to-R: S, UM, HM). As for body shape, the HM daughters had lower BMI (i.e., were slimmer) and lower waist-to-hip ratio (WHR, i.e., a more hourglass figure) than the other two groups. The authors make it clear that theirs is a correlational study that remains agnostic on the causal mechanisms, though Dienekes' post & comments discuss possibilities.
First, though, there's an interesting wrinkle: the authors and some of their references argue that the independent variable is presence of father (or parental cohesion), which yields the order S -> UM -> HM. The dependent variables are the markers of attractiveness, etc. However, the functions do not always turn out to be monotonic -- some curves decrease and switch to increasing like a U. Table 2 in the article summarizes the rank-ordering on the 3 facial markers. For "attractiveness," the order is UM -> S -> HM. For "health," it's UM -> S = HM. And for "masculinity," it's UM = S -> HM. Thus, if our graph has "parental cohesion" on the x-axis, the curve would be U-shaped for "attractiveness" and "health," though J-shaped for "masculinity." For body shape, Figure 2 shows that one variable, "impedence" (a measure of % body fat), wasn't significantly different among the groups. "Waist-to-chest ratio" (WCR) is a measure of the inverted-triangle shape of the upper body, something that men don't pay much attention to. The two important variables are BMI and WHR. On the latter, HM had significantly more hourglass figures than UM, though the S didn't differ significantly from either. On BMI, the HM were significantly slimmer than S, and apparently UM are in between. So, aside from perhaps BMI, the facial and bodily markers suggest that the underlying cause increases thusly: UM -> S -> HM. Call it "home harmony." The biological correlate of this in the literature the authors cite is response to stress (cortisol), so perhaps in the sample the daughters of UM parents experienced greater stress from the arguing, bickering, and so on, compared to the daughters of S parents, who at least weren't frequently fighting in the daughter's presence. Remember, the sample was of university students, so it likely didn't include those from the underclass or the lower end of the working class. That suggests that, above a certain threshold of SES, having antagonistic parents stay together produces more dissonance in the home than if they separated, at least from the daughter's p-o-v. Now on to the possible causes. Well, the first is what I just mentioned: the prevailing view that differences among father absent or father present homes reflect differences in stress during childhood. Humans have evolved a common set of responses, and those who happen to grow up without fathers turn out a different way from those who grow up with fathers. This is an environment / chance explanation. But as mentioned at the lead author's webpage and in the comments at Dienekes' post, there is also a (not mutually exclusive) genetic explanation: fathers who are apt to easily leave their wife & children, or who are too unruly for the wife to bother staying with, could be this way in part due to genes (perhaps for response to testosterone) which they pass on to their daughters. Moreover, since it's usually not impossible to read warning signs about who's less reliable & dependable than who else, we could also look at the mothers who mate with the more flight-prone males. Such females would likely show a greater propensity for risk-taking or thrill-seeking, presumably heritable, so assortative mating could be exacerbating the genetic influence of father. It would be interesting to take a large sample of males from S, UM, and HM parents and ask them who they were most attracted to among the S, UM, and HM female composite faces. That would settle whether there was an assortative mating effect. It would also be interesting to genotype those from S vs HM parents to see if the former were more likely than expectation to have the 7R allele at the DRD4 locus -- if so, that would suggest involvement of heritable personality traits like novelty-seeking, impulse control, and so on, in both the dissolution of the marriage as well as the suite of behavioral outcomes of the daughter. It might also suggest why the daughters of S parents were judged more attractive than those from UM parents -- presumably the S parents were more "wild child"-like than the UM parents who had to "wimp out" to some degree in suppressing their impulses to split up. Perhaps greater "wild child"-ness increases one's sexiness score (since "sexy" usually connotes something more exciting or thrilling than just "attractive" or "beautiful"). The prototype here would be Angelina Jolie, who looks more than a bit masculine, who's well known to be possessed of a thrill-seeking disposition, and whose parents divorced when she was a baby. To close, why would a tinge (though not an excess) of masculinity and rebelliousness make a female sexier, when these usually serve to make males sexier? These traits mix a "danger" component with the "beauty" component, which creates the thrill. My hypothesis is that, assuming the "cheesecake theory" of aesthetic pleasure popularized in How the Mind Works -- that art, cuisine, etc. are human devices to directly stimulate our evolved pleasure centers -- we enjoy stimulating not just the "relaxation" (or parasympathetic) division of our nervous system, but occasionally the "danger" (or sympathetic) division as well. Things that highly stimulate these divisions are, respectively, the beautiful and the sublime in an older terminology. Those whose aesthetic preferences lead them to want more than others to stimulate their "danger" system would appreciate a greater dosage of dangerous, masculine traits in females. Rawrrr.
More obviously non-controversial research on cognition and sex differences, Differences in cue use and spatial memory in men and women:
Science is like a box of chocolates, you don't get to know the details until you start rooting through it. Major tables with p values below the fold. 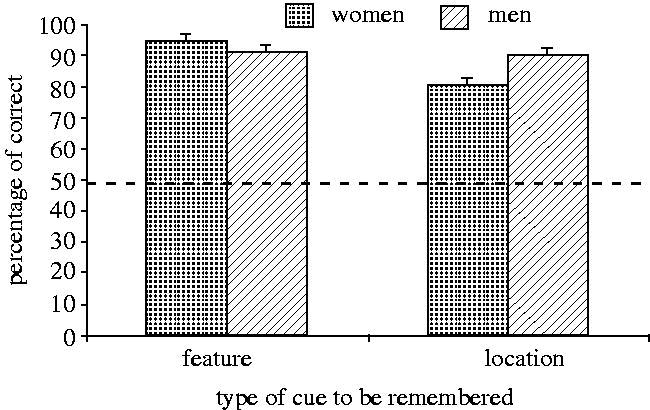 "Although the men tested tended to be older than the women (one-way ANOVA: F1,45=2.94, p=0.09) and the two experimenters tested subjects of significantly different ages (F1,45=9.24, p=0.004; experimenter A's subjects: mean, 22.44 years; range, 21-24; experimenter B's subjects: mean, 21.41 years; range, 20-24), age was not correlated with performance on either task (controlling for sex and experimenter; feature: F1,43=1.36, p=0.25; location: F1,43=0.45, p=0.50). Therefore, the observed differences in performance between the men and women on the location task is unlikely to have been due to differences in their ages." 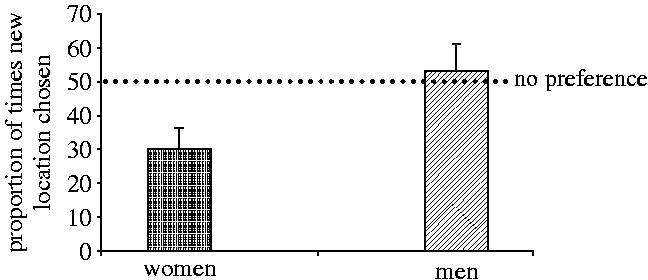 "One-sample t-tests were used to determine whether the number of times that the men and women chose the new location on the probe trials differed from random (i.e. a score of three). The choices of the men did not differ from random (n=20, t=0.42, p=0.68), but those of the women did (n=20, t=-3.33, p=0.004). Women chose the new location significantly less often than would be expected if they were making choices at random."
I've signed up for Clustrmaps before, but this weblog gets too many hits so it maxed out. This time I upgraded to the premium service and we shouldn't overtax it, so it is now there, above frappr.
A little while back, I mentioned epigenetics, noting that arrays for the detection of methylated sites in the genome are starting to become available. Recently released from The American Journal of Human Genetics is a new article that uses just that type of array to detect epigenetic variation in human sperm. what do they find? Here's the first paragraph of the discussion:
In this study, we performed an in-depth analysis to address the question of epigenetic variability in the germline. The main conclusions are that (1) the male germline exhibits locus-, cell-, and age-dependent DNA methylation differences and that (2) DNA methylation variation is significant across unrelated individuals, at a level that, by far, exceeds DNA sequence variation. That is, different people carry different epigenetic information in their germline. The next question is clear-- does this information get tranmitted to the next generation? In general, I had been under the impression that methylation was "cleaned" off the DNA after fertilization, but that's nothing but a general rule, and exceptions abound. If epigenetic information can be efficiently transmitted from parents to their children, that would be a huge--for the study of disease, for the study of genetic conflict, and for genetics in general. The authors of the article mention that the age of your father is a risk factor for schizophenia, and transgenerational effects on other traits have been seen before. Could this be the mechanism? This sort of phenomenon could also raise some hairy policy issues-- imagine that we know smoking causes specific epigenetic changes that, if transmitted to a child, increase their risk of some disease. Would we then be justified in banning smoking by anyone who is fertile? Or what if smoking decreases the risk of some disease in your offspring? Would the tobacco companies get back all the money they've paid out in settlements? Of course, I'm getting way ahead of myself-- no one knows how widespread the transmission of epigenetic variation will be. But this is fascinating stuff, and well worth keeping in mind...
Excellent review of the latest findings on sex influences and brain anatomy/functioning by Larry Cahill over at Nature Reviews - Neuroscience [open access]. One comment from Cahill:
A third, also related, misconception holds that the differences within a sex are much more substantial than those between the sexes, the implication being that sex influences can therefore be dismissed as trivial.Now, where have I heard this (sort of) argument before...?
Monday, May 29, 2006
I was pointed to this research (via David) that is just out about the correlation between variation on DRD4 and "sexual arousal." From the press release:
In short, it seems like a very sexual orientation is a derived trait, while the ancestral character tends to be more "repressed." My first thought was that someone should forward this to Geoffrey Miller, his theories relating to sexual selection and cognitive evolution are predicated on weak pair-bonding and operational polyandry as males and females form temporary relationships which dissolve within 5 years after sexual novelty has expired (you need lots of reproductive skew to really ramp up sexual selection, and polygyny is the normal way to go about that). In contrast, other theorists, like pervert-cum-anthropologist Desmond Morris, have posited the importance of pair bonding and monogamy in the natural history of our species. The relatively trivial sexual dimorphism among humans (around 10%) between the sexes in terms of overall size and canine ratio suggests a more monogamous past, while the sperm competition research implies a more "mixed" picture (one problem is that the words do not really map onto to the full distribution well). The short of it is that monogamous species tend to be less "sexy" in many ways because they don't need to be, whether that be in behavior, display or semen volume (much of this costs resources and so can detract from fitness). The spread of a sexy variant within the last 50,000 years is very interesting, because it is within the last 50,000 years that the cultural explosion has erupted that made us more than just "anatomically modern" humans, that is, behavorially modern. Honestly, I would have guessed that the sexy variant was ancestral, but this data will surely result in new stories being told over the next few years if future research confirms and elaborates on it, because sex + evolution = mucho $$$ in terms of book sales. The paper to published in Molecular Psychiatry is Polymorphisms in the dopamine D4 receptor gene (DRD4) contribute to individual differences in human sexual behavior: desire, arousal and sexual function. Click through and you will see why Jews are relevant, the study was done in Israel on university students, and found that particular DRD4 variants correlated with questionnaire responses. Is this generalizable to other populations? Well, DRD4 has shown up elsewhere. For example, DRD4 promoter SNPs and gender effects on Extraversion in African Americans. And of course, there is the Harpending and Cochran review in PNAS, In our genes, which argues that the variation in the allele frequencies between populations is suggestive of their respective evolutionary histories. I had some issues with figuring out their notation for the mutation, and the fact that I don't have full access to the journal article in question doesn't help, but here is a map from Alfred which shows regional variation at the promoter C/T mutation. On visual inspection I really don't have anything much to say in regards to the map and the allele frequencies (I assume that the C variant is associated with sexiness since it is in the minority). If any readers have access to the article I would suggest you might clear up of the extra-Israeli distributions of these variants as I wouldn't be surprised if it is mentioned in the discussion. Addendum: Perhaps sexiness evolved to "spice" up the monogamous pair bond?
The American Journal of Human Genetics has a paper in its pre-print section titled "A geographically explicit genetic model of worldwide human settlement history." I quickly skimmed it (and uploaded it into the GNXP forum). I have serious issues some of the inferences made in regards to the "obvious" fit of such coalescence data with a particular demographic history. I am convinced that meta-population dynamics tend to be ignored (in part because they are just another complication) even though they can also explain the data. Nevertheless, this jumped out at me:
As I stated above, meta-population dynamics, local extinctions and recolonizations, are issues that the authors seem to ignore when it comes to ignore their there results, especially given that environmental parameters are likely to be very relevant to marginal groups. They even allude to what seems like an abortive extra-African colonization in the Levant by anatomically modern H. sapiens sapiens which ended in local population extinction. But, the idea that humans are relatively buffered from environmental variation is roughly correct, and I'm talking about "humans" in a very broad sense. Our genus, Homo, broke out of the African continent almost immediately after its genesis. Dmanisi shows that beyond a shadow of a doubt.
Thought this might be of interest: Brandon Berg, one of my co-conspirators at Catallarchy, wrote a post a while ago sketching out a possible way in which the decline in fertility rates might be overstated in the statistics that are commonly bandied about. The basic idea is that if the mean age of motherhood is increasing each year, the stats are likely to be skewed because of the way they're calculated (simple summing of the fertility rates for each age group). Now he's written a short follow-up demonstrating how sensitive the fertility numbers are to this effect. He suggests that the European fertility rates could realistically be understated by about 0.1-0.3, which is not huge but still significant.
Update: Related: Population Fallacies Part 2
A new study tracks the effects of alcohol drinking during each trimester of pregnancy for 636 mother-child pairs.
Light to moderate drinking during the second trimester led to the most pronounced cognitive deficits at age 10. The catch is the effect was found only for the African-Americans in the study and not for the white children, suggesting that race differences may make alcohol drinking more harmful for the babies of African-American mothers: No such association was found for Caucasian children in the study. "This racial difference could not be explained by the amount or pattern of drinking during pregnancy or socioeconomic factors," [study chief Dr. Jennifer A.] Willford told Reuters Health. This suggests that genetics play a role in these racial differences, the investigators add. Also: "Our study also showed that prenatal alcohol exposure was associated with lower IQ for African-American but not Caucasian children, said Willford. "Importantly, we know that this racial difference was not due to differences in the amount or pattern of alcohol use during pregnancy or by differences in socioeconomic status. We cannot say why the racial difference exists, but laboratory animal and human studies show that it may be partly explained by genetic factors." Levitt and Fryer type sophism using motor development scales aside, cognitive differences between African American and white children show up very early, and persist in a number of transracial adoption studies (I'm collecting this research into an Ebook that I'll release here on gnxp when it can be finished). So studies like this are at least getting warmer than "acting white" type mythologies.
Sunday, May 28, 2006
The majority of cells in the nervous system are not neurons. They are glia. There are several subtypes of glia, one of which is astrocytes. Glia means glue. In general, people think of glia in their support role in the nervous system. Holding it together, disposing of waste and other menial tasks.
This little commentary here (pdf) in the current Cell describes a change in thinking that is occuring with regard to glia. They are stepping into a more active role. In this case, they modulate the ability of hypothalamic synapses to undergo plasticity. Compared to lactating rats, virgin rats have closer astrocyte encasement of synapses. The astrocytes release a chemical called D-serine into synapses that binds to a co-agonist site on NMDA receptors. NMDA receptor activation is considered the first checkpoint for increases in synaptic strength. More D-serine = more effective NMDA receptor activation. In the study described, virgin rats have a lower threshold for LTP induction than do lactating rats. This is nicely illustrated below the fold. 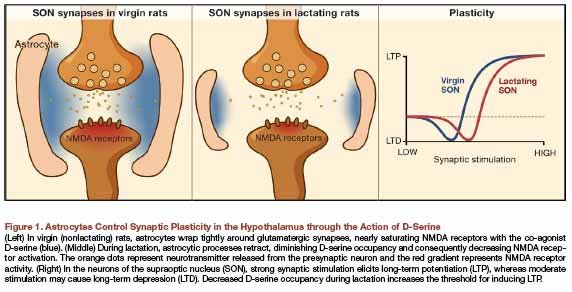 So better glia encapsulation of synapses = more D-serine in synapses = more NMDAR activation = easier to undergo synaptic enhancement. This is in the hypothalamus mind you, not usually a major locus for memory studies. I'd like to see what's going on during these states in the hippocampus. For those of you interested in intelligence and Ashkenazis and all that business, you might note that the major abnormality of Einstein's brain was an greater glia to neuron ratio. I don't know what that means either.
Saturday, May 27, 2006
Today's London Sunday Times has a good opinion piece by Simon Jenkins on global warming, here. My attitude is that we should leave our descendants to deal with their own problems, as they will be richer and better-informed than we are. Let us take decisions according to our own needs and priorities. But then I'm biased, as I've always disliked the 'doom-and-gloom' brigade, many of whom have a not-very-hidden anti-capitalist agenda.
This weeks Nature has a news article called "What is a gene?". Here's what they have to say:
In classical genetics, a gene was an abstract concept - a unit of inheritance that ferried a characteristic from parent to child. As biochemistry came into its own, those characteristics were associated with enzymes or proteins, one for each gene. And with the advent of molecular biology, genes became real, physical things - sequences of DNA which when converted into strands of so-called messenger RNA could be used as the basis for building their associated protein piece by piece. The great coiled DNA molecules of the chromosomes were seen as long strings on which gene sequences sat like discrete beads. This picture is still the working model for many scientists. But those at the forefront of genetic research see it as increasingly old-fashioned - a crude approximation that, at best, hides fascinating new complexities and, at worst, blinds its users to useful new paths of enquiry. ... The one gene, one protein idea is coming under particular assault from researchers who are comprehensively extracting and analysing the RNA messages, or transcripts, manufactured by genomes, including the human and mouse genome. Researchers led by Thomas Gingeras at the company Affymetrix in Santa Clara, California, for example, recently studied all the transcripts from ten chromosomes across eight human cell lines and worked out precisely where on the chromosomes each of the transcripts came from. The picture these studies paint is one of mind-boggling complexity. Instead of discrete genes dutifully mass- producing identical RNA transcripts, a teeming mass of transcription converts many segments of the genome into multiple RNA ribbons of differing lengths. These ribbons can be generated from both strands of DNA, rather than from just one as was conventionally thought. Some of these transcripts come from regions of DNA previously identified as holding protein-coding genes. But many do not. "It's somewhat revolutionary," says Gingeras's colleague Phillip Kapranov. "We've come to the realization that the genome is full of overlapping transcripts." ... Many scientists are now starting to think that the descriptions of proteins encoded in DNA know no borders — that each sequence reaches into the next and beyond. This idea will be one of the central points to emerge from the ENCODE project when its results are published later this year. Remember, when this blows up, you heard it here first!
Friday, May 26, 2006
Willing to Do the Math: An Interview with David Botstein on PLoS Genetics
Thursday, May 25, 2006
To interrupt your regular science programming, check out the back story on Miss "Poland." Turns out she is a washed-up Miss Venezuela, and that the reputed powerhouse of beauty contests has done some exporting before. Here is a gallery of the contestants for Miss Poland this year (not Kingdom of Saudi Arabia work safe, but since "work" in the KSA means letting some brown dude from brownland do everything I guess that's a dumb caveat). Hey, at least they didn't have to import from Japan to fill their babe quota because of a depauperate local environment :) (hat tip to Jaakkeli for the last link, who is associated with gnxp on the 6 out of the top 10 hits on google for his name)
Deadsmith dropped me another gem about a transcriptional profiling study in yeast getting a little more sugar. I will continue to share these with you until somebody tells me to stop. As before, any ire or accolades should be directed to him, as much as you can direct either one at a floating handle in cyberspace:
I really like the idea of studying things like cancer and Alzheimer's in things like yeast. Yeast are just so easy to do genetics on. I like things that are reliable and easy to work on (I also like 20 year old Toyotas). Okay, just file the following away... I may not even come back to it today: Despite the canonized pathways of metabolism in eukaryotes, notably yeast, there is still quite a bit about metabolism that is unknown. Metabolism is important for lots of things in biology, since biological organisms have energy requirements, but one particularly interesting aspect of metabolism for humans is how it changes in cancerous tissue. One of the hallmarks of tumor cells is the excessive metabolism of glucose, often the result of mutations that have left the glycolysis machinery running unattended. There. Now onto another topic: transcriptional response to environmental changes in yeast. Organisms, even unicellular ones, have to respond to their environment. They can do this in the short term by leveraging the proteins that comprise the cell against the problem (think heat shock proteins refolding proteins, for example). But if the stimulus is long-lasting, then the cell too needs a long-lasting response, and we generally start thinking about transcription of genes to make new RNA to, in turn, make new proteins to come to the aid of the cell in its hour(s) of need. But the details of these responses have not been particularly well-studied, so today I review an attempt to perturb yeast cultures while monitoring their transcriptional and metabolic responses (pdf) that was published by the Botstein lab at Princeton University's Lewis-Sigler Institute for Integrative Genomics in collaboration with Michal Ronen, a graduate student-turned post-doc at Stanford. Botstein has long been tangled in the development of microarray technology and interpretation, so it's no surprise that this study teams the microarray with an old microbiology tool, the chemostat, to profile the transcriptional response of yeast to small doses of glucose. The chemostat, for those unfamiliar, is a device that allows the steady-state growth of a microbial culture by continually adding nutrients and removing excess culture and byproduct. On a side note, the chemostat was invented by nuclear physicist Leo Szilard, Einstein's coauthor of the letter that kicked off the Manhattan Project (man, those guys all had night jobs!). Anyway, the idea here is to grow yeast in a chemostat, and then give them a tiny little glob of glucose to metabolize, and see how they respond transcriptionally and metabolically. The transcriptional measurement was done by microarray, and the metabolic assay was as simple as looking at how long it took for the glucose to "go away" and the ethanol to show up. I say "go away" because there are two obvious ways for it to do so: 1) glucose could be dissipating through the chemostat, or 2) it could be metabolized by the yeast. Since it "goes away" much faster if there are yeast in the chemostat, and since ethanol shows up on the chemostat if yeast are there, it's safe to assume that the yeast are metabolizing the glucose to ethanol, as yeast are wont to do. The interesting data, of course, is in the transcriptional response. The glucose doses came in one of two sizes: really small, and small. Both doses were too small to allow the cells to change in number (division) or size (growth without division), yet over the course of the next 3 hours (about one yeast cell cycle), 25% of genes changed expression at least 2 fold. Within minutes, there were dramatic expression differences, exhibiting the transcriptional response to the new carbon source, and over the remainder of the 3 hours measured, the RNA levels returned to their initial state in different modes. Most of the genes behaved in a "burst response," meaning that their transcripts dropped dramatically, then climbed back to initial state levels. These included the galactose metabolism genes, TCA cycle genes, and interestingly, ribosomal genes. The glucose metabolism genes, including both glycolysis and gluconeogenesis, both showed a "lasting response," though in opposite directions (glycolysis goes up and falls off slowly, gluconeogeneis goes down and comes up slowly). A couple of genes, including the hexokinase genes, exhibited a bidirectional response, first dipping quickly, and then peaking well above baseline. These guys are known to work in a cross-regulatory circuit with MIG1 and RGT1. MIG1 is responsible for repressing the expression of gluconeogenesis and galactose genes, and is upregulated by glucose presence in the cell (all this is outlined in the diagram I’ve taken from pg. 391 of the article and posted below). RGT1, meanwhile, represses the hexokinase transporter, which is responsible for importing glucose into the cell. The point is, these genes are in a repression-oscillator circuit, and thus have a bidirectional response that generates a smooth combined effect for regulating the switch from glucose to alternative carbon sources. 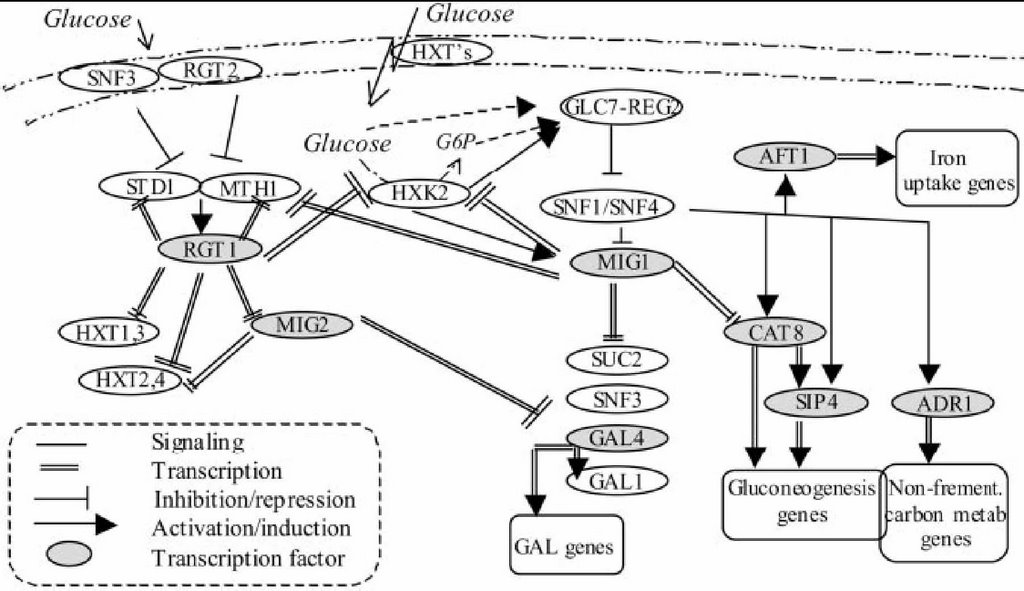 They also quantified the iron chelators, but I don't really see the significance, and the signal was much shakier, so I'll say only that about them. It's interesting to think about the transcript levels that fell quickly, as I went into this paper thinking that it was going to be about transcription, not degradation. The authors mention in the discussion that the half-life of a glycolysis RNA 3 times longer in a glucose-rich culture, and that begs for more investigation (since we also don't know jack about RNA degradation)[Coffee Mug note: we know a little about RNA degradation]. Using all this data on the levels of the RNA rise and fall of each group of genes, and knowing a bit about how each gene in the system regulates the system, the authors then construct a couple of basic differential equations to model the transcription factor activity (TFA) of some of the genes. It's important to realize that TFA is hard to measure directly, so if you can construct a good model of how you think it's working from the expression data, then you can create predictive groups of TFA by functionality. They did such grouping by functional annotations from the Gene Ontology database, and thus the end result was a set of mathematical classifiers for the types of transcription factor activity. As with all things yeast, this looks like a model study for something bigger. There are tons of other ways to perturb yeast, but it might be possible to look at these basic sugar metabolism circuits and their regulatory profiles (and predictive transcription factor activity models) and look at other cell lines. Maybe human cell lines? Maybe human cell lines with abnormal TFA profiles? Gee, what kinda human cells have abnormal transcription factor activity? Any with abnormal TFA that involve glucose metabolism that we care about? Hrmmm... It turns out that Botstein is interested in human disease, and I doubt this has escaped his attention, since I've seen him lecture on other ideas that involve studying cancer in yeast.
Peep the structure of a whole empire. - Malik B
I have a couple of nice papers here looking at the inter-relation between LTP, actin polymerization, and dendritic spine structure. They are both pretty dense, so I'll do a single post on this Fukazawa et al. paper (pdf) and then, with the conceptual junk already on board, its should be easy to present the information form this later study by Okamoto et al. The majority of excitatory synaptic transmission in the hippocampus occurs at dendritic spines. There appear to be specialized structural components on both the sending (pre-synaptic active zone) and receiving (spine, post-synaptic density) sides of a synapse that mediate efficient transmission. Modification of this structure provides a highly plausible mechanism for enhancing or diminishing the connection between two neurons in a lasting way. There is an intimate relationship between the structure of the spine and actin polymerization within the spine. Actin is a cytoskeletal protein with the ability to form long filaments to which other proteins can bind and interact in complexes. It is really remarkable that actin forms in integral part of synaptic protein complexes and helps to determine structure considering that the actual subunits that make up an actin filament turnover really quickly. According to Fukazawa et al., over 80 percent of actin in dendritic spines turns over in less than a minute. This makes one have to shift perspective a little bit and deal with actin as a dynamic process. All of the things actin does when it forms filaments must be regulated by altering the rate of polymerization or depolymerization rather than by a single polymerization event that is maintained once organized. Fukazawa et al. suggest a number of ways in which LTP might be realized through actin structure modification:
If this were a journal club talk I would be enjoined to get to the data already, but I have a couple more things you have to understand to get the paper, and I think it is interesting so you have to bear with me. In this Fukazawa et al. paper the LTP they are studying is actually done in a live animal. Most of the experiments involve inducing LTP via a specific pathway, and then staining the hippocampus to determine the state of actin. Actin can is found in two forms, globular (G-actin) and filamentous (F-actin). Death cap mushrooms make a poison called phallotoxin or phalloidin that binds to F-actin and not G-actin. The authors have phalloidin conjugated to a fluorescent molecule, so they can stain hippocampi and look for F-actin, indicating an increase in polymerization. The other cool thing is the specificity of the pathways they are stimulating. LTP was induced either by stimulation of the medial (MPP) or lateral perforant path (LPP) which project from the medial (MEC) and lateral entorhinal cortices (LEC) respectively. Others have shown that the MEC is specialized for representing spatial information while the LEC is more responsible for the non-spatial input into the hippocampus. The potentials generated in the hippocampus by these inputs generate different types of traces. LPP potentials tend to take longer to peak and last longer. The LPP and MPP both synapse in the same subregion of the hippocampus, the dentate gyrus, but they synapse on different parts of the dendrites of dentate gyrus cells (the outer molecular layer (OML) and middle molecular layer (MML) respectively, shown below. The LPP input synapses on dendrites further away from the cell body than does the MPP input. I find this intriguing because I also recently read that spines far away from the soma have thinner necks and are less able to diffuse calcium that comes in through NMDA receptors. As we all know, calcium starts doing interesting things if it hangs out in a dendritic spine very long. On the other hand, where does the calcium that diffuses through a stubby-necked spine go…into the dendrite where it could influence protein synthesis? This generates all sorts of interesting ideas about how modulation of spatial input could influence plasticity at non-spatial synapses. Would spatial information be remembered in a longer-lasting but fuzzier manner than non-spatial information? Or would calcium influx due to spatial representation modulate the ability of dendritic potentials further away from the soma to induce action potentials?  Back to the paper. The initial set of experiments involved inducing LTP just through the MPP to alter the synapses in the middle molecular layer (MML) of the dentate gyrus on one side of the brain and looking at what happens to actin and spine structure. Here is just one picture of a hippocampus labeled with fluorescent phalloidin:  Note that things are much brighter particularly in the MML. The level of F-actin has increased in the input layer has increased 45 minutes after LTP induction. They go on to show that this increase is happening in dendritic spines. In parallel, the average length of synaptic apposition (basically how much contact between axon and spine) increases in the stimulated side of the hippocampus. One thing to note is that, at least at this early timepoint, the number of spines does not appear to change. Rather it is the structure of existing spines that is altered by LTP induction. LTP can be induced to varying degrees with more or less high-frequency stimulation (HFS). In vivo LTP can last several weeks if induced with enough juice to begin with. Fukazawa et al. have short (HFS 90) and long-term (HFS 500) plasticity inducing protocols similar to E-LTP vs. L-LTP or LTP 1 vs. LTP 2 etc etc.. HFS 90 produces LTP that lasts around a day. The F-actin increase is observed 45 minutes after HFS 90, but not 1 week later. HFS 500 produces LTP and an F-actin increase that are still going strong a week later. Actually the F-actin increase is still there five weeks later, which is a substantial portion of a rat lifespan. The initial increase in F-actin is NMDA receptor dependent. For long-lasting changes in synaptic strength, actin must be allowed to continually polymerize at a higher rate (or depolymerize at a slower rate). This change in polymerization rate is in part mediated by synthesis of new proteins since cycloheximide (a protein synthesis inhibitor) and latrunculin A (an actin polymerization inhibitor) have the same effect of allowing LTP that lasts up to 8 hours but blocking any longer-lasting changes. The understanding of this process goes even deeper in this paper. The authors identify a particular promoter of actin depolymerization named ADF-cofilin that is negatively regulated by phosphorylation by a kinase called LIMK-1. That is, more LIMK-1 activity equals more actin polymerization (through a double-negative of regulation steps). The authors show that ADF-cofilin phosphorylation is higher in the stimulated side, and blocking this phosphorylation shortens the length of LTP. I find this last little bit especially interesting because it allows me to integrate some information from the first paper showing microRNA regulation of neuronal processes in mammals. MicroRNAs are recently discovered short bits of non-coding RNA that specifically inhibit the translation (either through degradation or sequestration) of complementary RNAs. The importance of miRNAs in gene regulation is equal to or greater than that of transcription factors. The mRNA for LIMK-1 is inhibited by a dendritically localized miRNA called miR-134. Plasticity inducing signals release the LIMK-1 mRNA from inhibition so that LIMK-1 can be synthesized. It seems perfectly plausible to me that this one of the proteins that needs to be synthesized to produce long-lasting LTP. Synaptic stimulation would remove miR-134 from the picture. LIMK-1 is translated. It phosphorylates ADF-cofilin. Actin is allowed to polymerize at a higher setpoint. Spines grow and synaptic contact is increased. So the story goes. It is probably more complicated than I made it out to be, but I will go into some of the problems with interpreting this work when I do the Okamoto et al. paper. One issue is that actin is all over the cell, including the pre-synaptic compartment, so it is difficult to perform manipulations that specifically affect dendritic spines.
The Roman Catholic Church's official policy on the 'regulation of birth' is, of course, that artificial birth control should not be used since the practice transgresses God's 'natural moral law' to go forth and multiply (and, of course, with congregation numbers already being as low as they are, well...). Therefore, "direct interruption of the generative process already begun...[is] absolutely excluded as lawful means of regulating the number of children."
The one loop-hole, however, is the rhythm method -- refraining from sex during a woman's fertile period -- one of those handy laws of nature which "God has wisely ordered." If you and your wife/husband just make sure to have sex only at those times when she's/you're less likely to get pregnant, then you can have your cake and eat it, too -- avoid unwanted prenancies, while not interfering with conception or having to opt for a selective abortion. (The rhythm method, by the way, is estimated to be anywhere from 75-90% effective in avoiding unwanted pregnancies.) However, in a forthcoming article in the Journal of Medical Ethics, Luc Bovens suggests that the rhythm method may work not (only) because it prevents conception from happening in the first place, but rather because those embryos conceived during a woman's less fertile period are actually less viable. Both the sperm and the egg may be old and the uterine lining may not be optimal. In the article [not yet available], Bovens offers some estimates of how many naturally aborted embryos we might be talking about -- the upshot is that he reckons that for every one unplanned pregnancy in rhythm method users in ten woman-years, there would also be two to three embryonic deaths. Even though the actual numbers can be disputed, the logic of the argument seems pretty sound (Bovens, via email): "If there is reduced viability for embryos conceived outside the abstinence period for rhythm method users, then the method is likely to be effective, not only because it reduces the chance of conception, but also because embryos so conceived have a lower viability. And hence for every failure of the rhythm method (i.e. for every pregnancy) there is a higher rate of embryonic death."If Bovens is right -- and if (to paraphrase) every embryo is indeed sacred -- rhythm method users may need to re-evaluate their 'moral choice'.
John Hawks added his two cents about noise including an interesting discussion on the source of circadian rhythms. Note that the real author of the noise post was not me (Coffee Mug), but a friend of mine who goes by deadsmith, when he goes by internet names..
Wednesday, May 24, 2006
In my recent post, A DNA database nation?, I commented that including everyone in a criminal DNA database would solve several ethical concerns with their current and expected use.
You can get a sense of the utility and concerns about DNA databases by reading the latest issue of Genomics, Society and Policy, which is apparently open to public access. Articles cover a number of topics: * Should police be able to access DNA samples collected for other purposes? * The importance of taking DNA samples from seemingly 'trivial' crimes -- many violent offenders had an earlier arrest for non-violent crimes * The inclusion of children in the British DNA database * The expansion of DNA profiling from the mere matching of samples with people to the prediction of race, physical traits, and behavioral traits from DNA samples * Genetics of criminal behavior * An interesting case study of DNA profiling from Norway I'll conclude by pointing out that most of these concerns could be circumvented if the DNA database was universal with sample collection delinked from criminal behavior.
Tuesday, May 23, 2006
Yeah, I never thought I'd write something with that title either, but serendipity happens. Today I read an article on mathematical cognition in adults and 4 year-olds from the new PLoS Biology, and I was trying to think of something neat to say about it, rather than just summarize what they found. So I put that on the back burner and decided to kill some time over at Edge, where this new vignette on happiness by Daniel Gilbert caught my attention. This quote from an interview on his website jumped out: as for the advice that you should buy the worst house in the hottest neighborhood rather than the best house in a modest area -- "It will make you feel bad because the brain is a difference detector; almost everything that it senses, it senses as a comparison."
Now, elsewhere he complains that psychologists suffer from physics envy, but lower-level research is necessary if we want a fully integrated picture. And sure enough, the authors of that PLoS article have shown that the Intraparietal Sulcus (IPS) is likely the locus of non-symbolic numerical processing -- that is, being able to tell how many things are in a visual array or how many tones are in an auditory sequence ("non-symbolic" because no numerals like 1 or 9 are involved). In brief, in both adults and 4-y.o. kids, this region of the brain lit up more when the number of circles in a series of slides abruptly changed from, say, 16 to 8 or 24. A different region lit up more when the circles abruptly changed to squares or triangles. The authors note that this did not involve mentally counting the items "one, two, three... sixteen... twenty-four," as many of the 4-y.o.s could not count that high. Rather, it must be the impression that "there are greater or fewer things than before." The circles in the slideshow varied in density, cumulative surface area, spatial arrangment, and size, so none of these could have formed the consistent "background" that the novel stimulus popped out of. The only sudden change was in number (or in the other experiment, shape). They cite other experiments that show that the mind intuitively processes such arrays, and presumably quantities of things generally, using a ratio rather than tallying the absolute value of each -- it's easier to discriminate sets with 6 vs 9 items than sets with 4 vs 5 items, since the ratio is more lopsided in the former case (deviates farther from 1.0), despite involving greater quantities in both sets compared to the latter case. In reviewing another study that didn't find the link between the IPS and non-symbolic numerical processing, they note that it didn't vary the surface area covered by the items -- so, the activation of the IPS there may well be due to detecting differences in surface area. This leads the authors to speculate that the IPS is involved in processing magnitudes in general, again using a ratio. So, if humans have a deeply rooted cognitive bias to ignore absolute value and focus just on the ratio between two things (or sets of things) that differ in magnitude, that would explain why having the worst house in the swankiest nabe won't make you as happy as having the most luxurious house on a street otherwise lined with run-down eyesores. Unable to appreciate the last-place mansion's absolute value, which most people can only hope for, we can only notice the lopsided ratio of its value to the mean value for the neighborhood. Sure, there's someone out there living in a delapidated tenement, but occasional, fleeting imaginations aren't as vivid and reliable as persistent realities. I commute to work at a tutoring center in a well-to-do Maryland suburb of DC, and I'm always amazed at how spoiled some of the kids are. But if brat A is only human, they can't fathom the absolute value of their luxuries, only how their stack compares to that of brat B. These observations aren't terribly new (i.e., that happiness is based on comparisons to others, and pie charts or bar graphs convey data better than raw tables), but they've been more fleshed out empirically -- we even know where in the brain one of the culprits is located -- and it's cool to see how seemingly disparate parts fit together.
I've been meaning to write this up for awhile now, so here goes. It's more of a pointer to what books & articles to read, so it may not cover every question you have about g and creativity. [1]
In Carroll's (1993) encyclopedia of cognitive abilities, Ch. 10 surveys psychometric data on the ability of "idea production" -- basically, creative thinking. It's a 2nd-stratum factor (called Gr for "general retrieval" ability), dominated by the 3rd-stratum general factor (g), thus a sister of the more well known visuospatial and verbal factors. Two of the 1st-stratum daughter factors that compose Gr are called FA (Associational Fluency) and FO (Originality/Creativity), which respectively measure how well you can draw associations between items in long-term memory (e.g., how interconnected your idea-web is), and how well you can devise novel solutions to problems (e.g., take ordinary objects & fashion new uses for them). Real-life tasks will also tap into Gc, or "crystallized" intelligence (the knowledge acquired as a result of applying g in academic / intellectual domains, e.g., vocabulary items, scientific ideas, etc.). So, given two individuals whose Gr is the same, the one with a larger store of ideas will have a richer web of associations. Simonton (1999 Ch. 2 & 3; 2004 Ch. 3 & 5) argues convincingly that at root the creative process is like Darwinian selection: ideas are combined in blind variation, largely unconsciously; most of these combinations are useless, but a few show promise. These are then selected and elaborated on consciously, putting flesh on the inspiration. Simonton says that aside from the usual suspects of high intelligence (g) and a healthy store of ideas & facts to draw on (unspecified, but must mean Gc), another part of creativity is having a "flat" associative hierarchy -- connections between ideas criss-cross a lot, rather than respect a "steep" hierarchy like Linnaean classification schemes. This trait is essentially Gr, though he does not recognize Carroll's terminology nor that it is dominated by g. This idea is originally due to Mednick (1962), whose Remote Associates Test measures how fluent one is in making remote associations. Ex: given the words "rat," "cottage", and "blue," they are all remotely related to "cheese" (the answer). French, Price, & Ekstrom's (1963) test has a subsection for idea production, one of whose tasks requires the subjects to fill in blanks to complete similes: e.g., "She was as pale as..." This test is a measure of diverse cognitive abilities, not just creativity, reflecting the consensus that there is no cognitive ability that is not influenced by g, and that all 2nd-stratum abilities correlate with each other. See Gottfredson (2003), where she tears Robert Sternberg a new orifice for lazily trying to argue that "practical intelligence" is different from and not correlated with g, one prong of his Triarchic theory of intelligence. Now, a skeptic like Michael from 2blowhards could say: OK, sure, if you restrict "creative products" to those endeavors that demand braininess, by definition creative people will be brainy, whereas if you took a more liberal & inclusive view in which "creative products" included more popular forms where the cut-offs for g are more forgiving, this effect would shrink or vanish. So, is it possible to avoid this tautology? Yes: define "creative products" roughly as "forms which inspire a sense of awe or marvel in the beholder, as if such products were not thought possible to come from mere mortals." If that sounds too reverential, you can use more folksy language, but you get the idea. Call this the "Wow!" definition. It avoids any mention of cognitive ability, favoring only the ability of the work to inspire, rather than to provoke the beholder to complain, "Meh, my 3rd grade son coulda done that." It also cuts across disciplines: it captures why we find the Michelangelos & Beethovens more creative than illustrators & composers of advertising jingles, and the Newtons & Gausses more creative than "lab men" & those who report another example of a phenomenon with thousands of attested examples. It may just so happen that humans are more wowed by feats of cognitive difficulty, but we didn't build this in -- it's just a quirk of human psychology. Martians might well be wowed by similes such as "The printer paper was as white as white printer paper." And it really is the cognitive part that we're wowed by -- no one but a boor would claim that Stephen King is a creative genius or super-intelligent based on his popularity / best-selling status. Even those whose personal tastes lead them to prefer King to Goethe are not confused about who is more creative or intelligent. They simply feel that King speaks to their tastes more than that arty-farty stuff. So, rather than a creative / non-creative dichotomy, we have a spectrum of more or less creativity. The farther one moves toward the creative end, the higher the demands on g in general and Gr in particular; the more one moves toward the less original end, the more relaxed the demands, as one is no longer re-inventing the wheel. I should also amend the definition to include only those forms which more or less originate with the creator, rather than forms which result from the individual "following a script." Thus we exclude actors, orchestra members, and individuals who solve math homework problems by means of common algorithms (like long division), as they're fundamentally different from playwrights, composers, and mathematical discoverers / pioneers. We expect the latter to be smarter, while we don't expect the former to be smarter (which is not the same as expecting them to be dull!). Lastly, intelligence -- whether g, Gr, or anything else -- is certainly not sufficient for high status in creative fields. Beginning with Galton, researchers of genius have noted that the distribution of "eminence" (a proxy for genius, as there are no unrecognized geniuses) is not the normal bell curve that we know and love, but rather log-normal. The less mathematically inclined can see pictures of what this looks like here; the more quantitative can read a nice pdf here on its use in the sciences, which has an excellent log-normal version of the Galton board. The key difference is that, while a normal curve is symmetric, the log-normal curve is skewed; for eminence, it is highly skewed, like the green or pink curves on the NIST link above. At first, you might think: "Well, maybe that's just the far, far-right tail of a bell curve -- the super-duper nerds." But the shape is not the same. If you look at the "slope" of the right half of a bell curve, it changes from "steep" descent to "shallow" descent pretty quickly -- by the time you got to the far-right tail, you would only see a shallow descent. With the log-normal curve, however, you see a change from steep to shallow, so it can't be the same as the far-right tail of a bell curve. ("Steep" and "shallow" refer to the absolute value of the slope (rise / run) at a point: if it's greater than 1, i.e. "rising" more than it's "running", call it steep; if it's between 0 and 1, i.e. rising less than it's running, call it shallow.) So what's the big deal? Well, log-normal curves usually imply that there's a synergy among various components that produce the effect -- where the whole is greater than the sum of its parts. (Or: where the effects of the random variables involved are multiplicative rather than additive -- if the effects are additive, the result is also a normal bell curve.) To take a more concrete example, consider female facial beauty -- let's say there are 5 variables involved: eyes, lips, hair, prominence of bone structure, and rough facial geometry (e.g., long or short face). Assume there is a universal scale of less-to-more beautiful values for each variable (e.g., fuller vs thinner lips, larger vs smaller eyes, etc.). Also assume each variable would yield a normal bell curve. Now, in an additive model, you would take the points earned from each variable and sum them up -- so, the lips variable doesn't "see" or "care about" the eyes variable; you get X points for full lips no matter what. By the Central Limit Theorem, the sum of these 5 variables would itself be a normal bell curve -- on a scale of 1-9, most people would be a 5, and the percentages would drop off in both directions, so that there would be very few 2s or 8s. That's the opposite of reality, though: the percentage of 2s is far greater than that of 8s! This is because the variables "see" or play off of each other -- large eyes and full lips produce a more beautiful effect than if you just added their values together. Likewise, large eyes, full lips, lustruous hair, and prominent cheekbones -- 4 great qualities -- can all be brought crashing down by a long horse-face, rather than only suffer a slight reduction in beauty. In the case of creativity, there are several ideas for what these other factors are that interact synergistically with intelligence -- e.g., the Big Five personality trait Openness, Eysenck's personality trait Psychoticism, schizotypal personality, and so on. This is another interesting area of creativity research, but it doesn't mean that intelligence is any less important as a result. Consequently, claims to the effect that "IQ doesn't matter in creative fields past a threshold of 120" are nonsense. It's hard to get large data sets for such situations since there are damn few Newtons and Beethovens alive to test. Moreover, let's say it turns out that when you do the multiplication of the variables for arts vs sciences, the intelligence variable is weighted more in the sciences than in the arts -- nevertheless, in any cognitively demanding area higher *g* always helps. [1] For more background on g, sub-g factors, and brain correlates of g, see these two posts from the GNXP archives. Refs: Carroll, J.B. (1993). _Human cognitive abilities: A survey of factor-analytic techniques_. Cambridge: CUP. French, J.W., Ekstrom, R.B., & Price, L.A. (1963). _Manual and kit of reference tests for cognitive factors_. Princeton, NJ: ETS. Gottfredson, L. S. (2003). Dissecting practical intelligence theory: Its claims and evidence. _Intelligence_, 31(4), 343-397. Mednick, S.A. (1962). The associative basis of the creative process. _Psychological Review_, 69, 220-32. Simonton, D. (1999). _Origins of Genius: Darwinian Perspectives on Creativity_. New York: OUP ------------- (2004). _Creativity in Science: Chance, Logic, Genius, and Zeitgeist_. Cambridge: CUP Addendum: In the comments people are bringing up what I hope they wouldn't bring up, since it's a whole 'nother post, but I saw it coming! So a word or two about racial differences in creativity. First, unlike the copious data on the 1-SD difference in the means between Af-Ams and whites, there is no similar data on tests only of Gr, so all we have is speculation. That said, my guess is that, while not doing so well on g, Af-Ams do pretty well on Gr -- lopsidedness in the 2nd-stratum factors isn't unheard of. On average, Af-Ams and Ashkenazi Jews have greater verbal than spatial sub-scores; NE Asians are the reverse. Commenters have mentioned hip-hop music, but that's the wrong place to look -- Jazz for sure. Also, the popular phenomenon of "yo momma" jokes is basically a modified remote associates / similes test. True, you can cheat by stealing someone else's joke, but word gets around fast, and unoriginal jokes are quickly booed. You can see all this play out on the new MTV show "Yo Momma." The more remote the assocation, while still making sense, the higher the score; ditto for verbal cleverness. For example: "Yo momma sweat butter and syrup and got a job at Denny's wiping pancakes across her forehead." "Yo breath smell so bad the only dis I'm gonna give you is dis-infectant." And so on. However, all of this is at the popular level, while what I was writing about was high culture -- so Jazz would still survive, but not the other stuff. If you want to include popular culture as well, these products don't inspire as much marvel in the beholder, so the cognitive difficulty (which is what really inspires awe) isn't as demanding. Thus, the g variable is weighted less, but remember: creativity results from multiplying together a host of factors. All I was arguing was that g was one of them, and likely heavily weighted. Some of these other factors may favor Af-Ams -- for example, people who reach high eminence in creative fields are usually more disagreeable than agreeable; at such heights, diplomacy is for suckers. Again, I'll take it for granted that the Af-Am mean is more in the disagreeable / confrontational direction than is the NE Asian mean. So that's in the Af-Ams' favor. But I also mentioned Openness to experience, and Af-Ams seem to be more conventional and less tolerant of novelty, fantasy, thrill-seeking, and so on (this difference is often the seed for black comics' jokes about how whacky white people are). On the plus side, that renders them more immune to New Age flimflam, yet it also is a penalty when we consider the factors involved in creativity. As for NE Asians, they have higher g on average, their strength coming from superior visuospatial skills. So, look at how good they are at innovating in visual areas of pop culture (their poor turn-out as comedians, etc. would be due to lower verbal skills). I've mentioned before that they're pretty innovative when it comes to visual tasks, provided they're financially secure: graphic design, hell, any design, film, video games, and so on. But as for the Openness, Agreeableness, and schizotypal / eccentric-nutty behavior, I'd guess they tend to score in the direction penalized in the creativity multiplication. I'm sure some of this is cultural, but still, exposure to & incorporation into mainstream American culture still leaves the (correct) stereotype that NE Asians are more conformist than whites or Af-Ams. Less Open and more Agreeable individuals will appear more conformist. To the extent that there is a genetic component to these personality traits, then there will be a ceiling that they'll hit even when the cultural obstacle is removed. These observations pertain as well to the male-female gap in creativity. This is particularly apropos given the Larry Summers fiasco. In order to show discrimination, you'd not only have to take into account the different variances in the male & female curves for g, but the potential differences in means and/or variances for all the other curves involved in the multiplication. Not only are males more likely to have an IQ of 155, but they're certainly more likely to be Disagreeable and schizotypal / eccentric (my guess: difference in means), as well as to be the more Open / daydreamer sort or exhibit nonconformity (my guess: difference in variances). If you look at the real loonies among scientists and artists -- mathematicians and composers -- they're so overwhelmingly male, the culprit can't be g only. In these fields, females are penalized not only for lower likelihood of reaching say IQ 155, but also for on average being more Agreeable & diplomatic, not Open enough (too practical), and not nutty or eccentric enough -- and remember, these penalties are multiplied or compounded, not merely added together. It's hard to avoid the obvious: at the high culture level, males of Eurasian origin (as in, west of the Himalayas / Siberia) dominate more than would be expected just based on g, though that certainly plays a role too. My hunch is that, for whatever obscure reasons, their average values for each of the variables involved in the multiplication result in greater creativity than other demographic groups. Cultural factors can dampen or amplify this pattern to an undetermined extent, but those are the differences we're starting with. When we move down to the popular, less awe-inspiring level of culture, where the demands on g are less intimidating, then other groups will make greater headway -- again, following the pattern of their average cognitive profile (more verbal for Af-Ams and Ashkenazis, more visual for NE Asians). Labels: IQ
The most recent Current Opinion in Genetics and Development has a number of articles grouped around the theme "The sex chromosomes and human disease". A bit that might be of interest from this one:
The accumulation of intelligence (i.e. MRX) genes on the X might have occurred as the result of sexual selection - the choice of smart males by choosy females. Given that sexual selection is extremely rapid, this could explain the huge enlargement of the hominid brain over the past million years. I'd heard the sexual selection argument for brain size before, but never in the context of genes for intelligence on the X chromosome. Apparently I should have, though, because they reference an article from 2001 as their source. Here's the original hypothesis: Therefore, regarding human evolution we propose the following model: one of the most important factors contributing to the uniqueness of human evolution is that at some point human females decided to select males according to their advanced cognitive abilities. The same cognitive abilities are selected for in the struggle for survival. In humans, the development of the mating characteristic is augmented by natural selection. This is an ongoing process with exponential acceleration, which will propel the development of general cognitive abilities in humans into areas we cannot imagine now I think everyone can agree that the last bit about "exponential acceleration...into areas we can't imagine" is laughable, but what about the rest? Anything there? I imagine natural selection acts quicker on X-linked genes than on autosomes anyways; it'll be interesting to see how people try to tease apart the two. UPDATES: 1. Razib had a thread about this on his other blog a little while back. 2. To answer the question in the comments about why they think this is sexual selection, the answer is pretty much that genes involved in reproduction are over-represented on the X as well. So, well, yeah. I'm not too convinced. 3. Interesting factoid: linkage studies don't usually look at the X. This one[pdf] certainly didn't. Something to ponder...
A few papers out right now, like Jackson and Rushton, revolving around Frey and Detterman's formulas for converting SAT scores into IQ scores. First, gnxp's own A.A. Beaujean has a paper in the latest Personality and Individual Differences, providing additional evidence that the SAT is a reasonable measure of g, matching it up reliably to another set of undergraduate scores; though it is suggested that the conversion equations need to be improved, which will take additional research samples. Second, the versatile evolutionary psychologist Satoshi Kanazawa provides yet more IQ and the Wealth of Nations-inspired work with his new, in-press paper IQ and the Wealth of States. (PDF here) Chapter 5 of IQ&tWoN set the foundations for the book's larger cross-national comparisons, by first showing that regional, sub-national IQ scores also predicted differences in economic prosperity between regions. For instance, within-city IQ differences between districts/boroughs in New York City and London in the 1930s were highly correlated with intra-city economic differences. Similarly, economic differences between a number of 1930s American cities showed a significant relationship with childhood IQ differences between these cities as well. A study of IQ differences between US states circa 1950 (using large-scale military data) found a correlation of .81 between IQ and state income. Other data is provided for regions of France, Britain and Spain.
Using Frey and Detterman's equations, Kanazawa now attempts to update the 1950s study by comparing converted state SAT scores with state economic indicators. But a simple comparison is complicated by the fact that SATs are not taken by representative samples of the population, but by an upwardly biased group of college-bound high-school graduates. Also the percent of people taking the SAT differs dramatically by state. Kanazawa attempts to mathematically "correct" for this and his success seems mixed. The results go in the predicted direction; for instance the correlation between the converted state IQ scores and the Gross State Product (GSP) per capita is .50, with median family income is .57, and with % in poverty is -.35. These are fairly high associations (higher than the relationship between IQ and individual economic success, in fact), but it's possible that the even higher 1950s state correlation is more accurate. The selection bias of the SAT is already missing at least one large part of the story - as Kanazawa notes, even unconverted state SAT scores do not correlate with % of the population that is black, despite the large, well-confirmed black-white IQ/SAT gap. He doesn't attempt to explain this, but it could mean the number of blacks actually taking the test is fairly similar in each state regardless of the total number of blacks in each state. Perhaps readers have a better understanding of what's going on. Another anomaly is that the reported state IQ levels themselves are implausible, which is probably a function of both the Detterman equation and the Kanazawa conversion - or to put it another way, the imperfection of converting the SAT into IQ (or SAT as IQ) or to adequately "correct" for the SAT selection bias. For instance, the highest state IQ is New Hampshire with an IQ of 110.3 and the lowest is Mississippi with an amazing 62.7! That's a spread of over 3 standard deviations - almost 50 IQ points. Fully 12 states are reported with IQs lower than 80, and this is with race not playing a detectable role. We get Utah with an IQ of 75.1 and Wisconsin with 78.4! We can compare this against other data to see if it's compatible. For instance Henry Harpending provided Steve Sailer with Project Talent IQ data for 366,000 high-schoolers from 1960. This representative data only has a spread of about 1 standard deviation: from Montana with an IQ of 105 to Alabama with 89. The Encyclopedia of intelligence also provides Wechsler standardization differences by U.S. region. (by lumped states) which gives the modest spread of 101.6 for the Northeast, to 98 for the South (3.6 point difference). Kanazawa's numbers lumped in the same way provide much different results, with regional differences exceeding 1 SD. Kanazawa is aware of all this and suggests: . . . while the state IQ estimates do correlate very highly with the macroeconomic performance measures and thus appear to have some validity, it is difficult to take the estimates at the face value. . . Until more accurate estimates of the absolute levels of state IQ appear (derived, for example, from actual IQ tests administered to large, representative samples of state populations), perhaps it is best to treat the current estimates as reflecting the relative standings of states. . . than estimating the absolute levels of state IQs. Finally, it was worth applying this information to the infamous blue state/red state IQ hoax from 2004, and Kanazawa's data were not kind to the red states. It matters not if we accept his relative or absolute scores, the states stack blue side up. In the link above, Steve Sailer showed only small differences on the NAEP achievement results, but with Kanazawa's SAT aptitude results we find that 8 of the top 10 scoring states were Gore voters in the 2000 election (average blue state IQ = 99.3) and 8 of the bottom 10 scoring states were Bush voters (average red state IQ = 90.2). So, apparently, those ornery hoaxers were on to something after all.
Monday, May 22, 2006
An old friend of mine (handle: deadsmith) dropped me this discussion of selection and noise in biological systems. I figured with the various discussions of noise around here, you folks might enjoy it. The words below are his, the errors in formatting are mine:
I’d love to tell you that this entry is going to be a comprehensive, cogent, and readily accessible one-stop shopping spot for all the engaging literature on noise in gene expression that has been burgeoning in the background of your copies of Nature and Science for the last ten years. But, that’s already been written, by a far more credible source in Jonathon Rasner and Erin O’Shea. So, I’m going to recap some of that review here, and try to wrap it gently in the cushy context of why I was interested in the first place. If you get interested in evolution on your own, it’s probably because you read a pop-science book by someone like Gould or Dawkins or Ridley; or maybe a monkey person like one of the Leakeys or Marc Hauser or some such. And if you do, then you’ll get told about the forces of evolution, namely: Selection, Drift, Flow, and noise. The last one doesn’t get a capital letter here, because it almost certainly didn’t get one in the books I’m talking about. And that’s the point: noise is last in the list, because selection is the most important force, followed by drift and flow, then noise is a distant factor. Right? And what is “noise” anyway? When reading the intro-to-evo books, it sounds a lot like “unexplained variance in the object of study.” And I think lots of people are still using it this way, even when they mean more specific concepts, such as “stochastic,” which is not the same thing as “unexplained variance.” And where does this noise come from? And why did it come from whence it came? Is it staying for dinner? I think we tend to think of biological organisms as elaborately crafted systems with relatively precise mechanisms, often analogous to engineered circuits, e.g. feedback loops, oscillators, etc. And if you’ve ever done any engineering, then you know that there are elaborate equations that engineers use to go about estimating and minimizing the noise in their systems, so why wouldn’t natural selection have done the same thing? Maybe furthering the engineering analogy can pursue the answer. You’ve probably purchased an electronic component at some point that failed prematurely, and you’ve probably purchased one that lasted longer than was expected. I won’t name any brand names here, but we all know that some companies spend more in quality assurance than others. Why? On one hand, some companies pick the strategy of spending a little more and making a quality product, knowing that if it’s important to the customer, then they can charge a little higher price for their product. This might be common in the automotive industry, where people don’t want to buy new cars every week, even if they cost $50 a pop. But there are other industries where things are more disposable, because people don’t need them everyday like many people need their cars, such as point-and-shoot cameras. Why cheap out on the camera? Because it’s expensive to pay engineers to do those equations, to machine the parts to higher specs, and to have a lower threshold for crappy components that get chunked in the dumpster out back before they ever hit the shelves. So, people won’t die if their cheap camera is taking discolored pictures nearly so much as if their wheel falls off on the interstate. Okay, enough analogy. Selection should minimize noise, but minimizing noise in gene expression, just as in electrical component failure, is a complicated venture that has to take into account all the factors of the entropic universe and might take quite a bit of extra machinery. So, if a little noise can be tolerated, then why not just make a crappy camera? Oh, evolution, you are a shrewd craftsperson indeed. And if you’ve made it down the lay-science best sellers list to people like Dennett, then you probably encountered the idea of an evolutionary “design space“ ‐ that is, a theoretical space for the course of new phenotypes to play in en route to possible selection, and a little noise might just broaden that space a bit, without being detrimental to the existing organism. Okay, really, enough analogies, let’s talk molecular biology. Single celled, monoclonal organisms are ideal for studying the noise in gene expression, because you can setup a reporter system and measure how much variation there is in the clonal population in the same environment. As all good... well, at least all... intro biology books tell us that the phenotype is the product of the genotype and the environment. A + B = phenotype. Did you see a noise term in that equation? I didn’t think so... Think again. It so happens that even with genetically identical cells, in an identical environment, we see variation in phenotype. I’m not talking twin studies here, so much as single cell organisms floating in the same pudding. One paradigmatic method in the contemporary noise papers comes from the Elowitz lab at CalTech (at least I think it started there). Elowitz noticed that some monoclonal cells weren’t behaving identically, even when they were in experimentally identical environments. So, he took some CFP (cyan fluorescent protein), and some YFP and stuck them both behind identical promoters in one case, and different promoters in other case. This allowed him to compare two reporters that either had the same promoter, thus the same transcriptional regulation (or at least a big part of it), with two reporters that had different promoters. From there, you can subdivide the noise into “extrinsic” and “intrinsic.” Extrinsic noise sources will result in the expression of each reporter changing similarly, and intrinsic noise affects the reporters differentially. So, you can think of the noise source from the perspective of a single reporter. The best example of an extrinsic noise source is probably cell cycle stage. If the clones are all in a different stage of their life cycle, then you would expect gene expression to vary considerably (this, by the way, is an example of using the word noise to mean “unexplained variance” rather than “stochastic variance”). If the cells were synchronized, then you can see if some of the expression variance from the same gene across many cells goes away. But if you’re looking at two reporters of the same gene (or at least the same promoter), in the same cell, then you can measure the intrinsic noise of that gene’s expression. Then you can combine the two fluorescent reporter construct with manipulations of gene regulation, e.g. varied transcriptional and translational efficiency. To vary transcription, you might mutate the promoter of the reporters or maybe use an inducible system with a variation in how much inducer you supply across conditions (think lac operon); to vary translation you could mutate the ribosome binding sequence or maybe the initiation codon (Ozbudak, et al 2002). Then you could see the relative contributions to noise of transcription or translation, and in the case of E. coli, it appears to be translation. The story behind this is that the prokaryotic translation process occurs simultaneously with continued transcription, and there is competition between the ribosome and RNases that want to eat the mRNA for access to the ribosome. This competition results in the ribosomes blocked from the mRNA, and RNases degrading the mRNA. So, you either get translated, or you get degraded, and if you assume a constant probability of translation, and that each event is independent of the next (and you’re a statshead), then you’ll recognize this as the geometric distribution and you could model this dynamic as McAdams and Arkin did in 1997 and predict what would later be confirmed by the Ozbudak study. And that’s what happens with the two-reporter system in coli, but what about eukaryotes? The lab of J.J. Collins looked into eukaryotes with the two-reporter system cloned into budding yeast (S. cerevisiae). Collins notes that in eukaryotes, transcription is conducted in pulses, or for your neurobiology people out there, in quanta. In contrast to the prokaryotes, which are expected to have difficulties translating their mRNAs once transcribed (due to the RNase competition mentioned above), the eukaryotes were predicted by the Collins lab to have troubles initiating transcription, and that transcription would be the major source of noise. When the initiation failed, there would be no mRNA, and when it succeeded, then there would be a stabilized promoter, allowing multiple transcripts to be reeled off the DNA. They induced the GAL1 promoter (native mechanism of yeast to produce an enzyme for galactose when the preferred sugar glucose is unavailable). Usually, the GAL1 promoter is activated by galactose, via its UAS (upstream activating sequence) and an activator responsive to galactose. Using this context, and also an artificial one employing a tet-O construct, the experimenters were able to poke and prod the transcription initiation of the gene. And whichever ways they went, the findings were that transcription was very noisy when the gene was moderately induced, but as the induction increase, the noise went down. Meanwhile, they used codon manipulations known to affect translation efficiency (this uses codon bias and the CAI for anyone interested, but I’m not going into it here ‐ suffice it to say, there’s a better way to do this experiment, but this worked well enough to fetch a Nature paper). The results: eukaryotes had noisy transcription, and translation that was robust to noise ‐ just the opposite of the E. coli bugs that Elowitz developed the two reporter system in. And just to make things complicated, the O’Shea lab (see above) took their own shot at eukaryotic expression noise using nearly the same system as Collins, but wanted to characterize intrinsic vs. extrinsic transcriptional noise. The results: it varies from promoter to promoter. It’s really a crime for me to only say this much for the O’Shea study, but as it’s really well written, I think it’s better extracurricular material for anyone that’s really interested. So, to recap: 1) prokaryotes have noisy translation because their transcripts are fought over by RNases and ribosomes. 2) Eukaryotes have noisier transcription because activation of promoters leads to pulses of mRNA production, but 3) this varies from promoter to promoter. Now... Why would it vary? This is how we come full circle. There are two rationales for having a noisy gene. First, it may not have been that important to refine the noisiness of the gene. It’s expensive to build in more precise control circuitry, and if you don’t need it, then you don’t need it ‐ sell the darn camera. On the other hand, what if it were a trait that didn’t need to be precisely regulated, thus was allowed to stick around and be noisy, thus provided variation in a phenotype amongst a monoclonal population? Would that give an advantage to the noisy population of clones in a time of stress? A time where the selection pressures were a little more specific? Is noise a way to allow organisms to hedge their bets when selection isn’t currently holding their feet to the fire on a particular issue? Many think so. But this isn’t just conjecture; there are hypotheses here. One that has been tested: Are essential genes more likely to be noisy than others? Using the stochastic models developed by the Collins lab, the Eisen lab at UC Berkeley computationally analyzed yeast genes and found that the noisier the gene expression, the less likely the gene was to be essential ‐ thus evolution isn’t willing to play around with the bread winners of the genome. What’s more? The same experimenters point out that whereas extrinsic noise changes gene expression, it changes all gene expression in the cell without regard to specific genes. Then they posit that genes that are not part of a multi-subunit complex would better tolerate this noise. Using data from two near-proteome-wide mass spectrometry studies, they show that proteins in complexes do appear to be less noisy than others. Hot damn. Molecular evidence that selection crafts adaptations to be more precise when it’s more important. And when you phrase it that way, maybe it doesn’t sound so revolutionary. But these researchers are showing us not just which genes are expected to be more carefully engineered by evolution, but precisely where the tradeoffs in the machine are ‐ and the tradeoff points are different in the prokaryote machine versus the eukaryote machine. What’s more? There are predictions about where to look for the next essential genes. All the genes that are already essential are tightly regulated, thus have less phenotypic variance to select from. Meanwhile, the noisy genes could be the accidental saviors of a species when the next heavy selection pressure pops up. I suspect that if you start mapping things like essentiality and multi-subunit complexes to molecular phylogenies, then you may see a trace of this in the natural history, and then you’d have a historical account of the lattermost evolutionary force that even the most selection-centric evolutionists might write about.
In less than two weeks GNXP will be four. A small little weblog that started out at 300 uniques per day in the summer of 2002 is now steady at 3,000 per day in 2006 (and, has spawned a spin off weblog). We've been #1 for gene expression on google for a while now, and have slipped on to the second page for the gene query. We've grown at a slower clip than the rest of the blogosphere over the past few years, but what we lack in quantity of readers I suspect we make up for in quality. In any case, I've bullied Jason Malloy into writing some sort of retrospective by June 2nd, since he remembers "the blog" before there was a blog (when Godless Capitalist and I had our own places on the web), and tends to have an encyclopedic knowledge of all things GNXP. If you have stories & anecdotes that might spice up the post, drop them in the comment boxes. Especially old commenters who have gone into hibernation (though people reemerge, like bbartlog).
Also, I'd like to welcome JP, Darth Quixote and MC Coffee Mug into the stable. For those of you who are a bit older, I imagine this is going to be a "triple threat" like Jordan, Pippen or Kukoc, or Bird, McHale and Parish, deep into the future.
At my other blog...I post about why The Da Vinci Code is important (unfortunately). Also, a quick introduction to David Haig's kinship theory hypothesis re: genomic imprinting.
Glove compartment, gotta get my cash. Cause the crooked cops’ll try to come up fast. - Chamillionaire
If you’re interested in the molecular mechanism of learning and memory, you read a lot about synaptic plasticity and long-term potentiation (LTP).. If you’ve read anything at all about LTP, you’ve probably read that there is this special molecule called the NMDA receptor that serves as a “coincidence detector,” and allows neurons that are firing at the same time to increase their connection. The NMDA receptor is an ion-channel that, when coincident firing is detected, allows calcium to flow into the cell. Calcium inside the cell becomes more than just a charged molecule. It acts as a second messenger, binding to several enzymes and increasing or decreasing their activity. The signal transduction cascades that follow can lead to structural and biochemical modifications that increase the response to neurotransmitter release onto the stimulated synapse. The NMDA receptor, however, is not the whole story. There are numerous sources of calcium within and around a cell. This Journal of Physiology paper describes three of these other sources and dissects the role of each in determining the duration of changes in synaptic strength. LTP in the hippocampal slice is the model of choice for folks trying to figure out mechanisms of memory at a molecular level. It can be observed in plenty of other preparations, but the hippocampal slice far and away the most common. I think one of the reasons is that the hippocampus is a brain region known to play a role in the type of memory we refer to day-to-day: episodic memory, so if we can figure out how connections are made there, then we are closer to having memory with a capital ‘M’ figured out. Another reason is because it’s relatively convenient. To study LTP you need to be able to stick a stimulation electrode in one area and a recording electrode in another area that receives that stimulation. The hippocampus has a stereotyped circuit diagram in which these connected areas are easy to spot. To induce LTP, you use test-pulses to get a baseline and then shoot some intense stimulation in. After the high intensity stimulation, the response to test-pulses over at the recording electrode end increases. The observed increase is LTP. There is more than one type of LTP. One of the major distinctions has been between potentiation that requires new protein synthesis (translation) and lasts several hours (Late-LTP / LTP 2) and that that can be achieved with the proteins lying around already and lasts about 90 minutes (Early-LTP / LTP 1). The authors here make a further distinction for LTP that lasts even longer and requires synthesis of new mRNAs (LTP 3). The three LTP-types are induced in order of duration by increasing induction stimulations. LTP 1 is induced by a 1TBS protocol (theta-burst stimulation), LTP 2 by 4TBS, and LTP 3 by 8TBS. The number before “TBS” indicates the number of “shots“ the stimulation electrode is giving. The graph below shows the results of different induction protocols.  Okay, so here’s the awesome part. The three induction protocols specifically activate three different calcium sources that correspond to three different cellular compartments. The authors use pharmacology and calcium imaging to isolate the individual contributions. The calcium imaging is achieved using low-affinity calcium binding molecules that fluoresce when they bind and really high-resolution microscopy (two-photon). Here are the attributes of the three sources:
Inositol triphosphate (IP3) can be generated by metabotropic glutamate receptors (mGluRs) situated more peripheral to the synapse that are probably stimulated only when enough neurotransmitter is released to spill-over past the AMPA and NMDA-type receptors in the central part of the synapse. The IP3 binds to IP3-receptors in the ER out in the dendrite and induces calcium release. The VDCCs do what they say they should. Once the membrane potential at the cell soma reaches a sufficient depolarization these channels will open up and let calcium flow in right there next to the soma where it is ideally positioned to activate transcription factors and initiate mRNA synthesis. Put this all together, and you get a model like the authors proposed here:  One further thing I’d like them to investigate if they got the chance is the synapse-specificity of the LTP 3 that is generated in the presence of an NMDA receptor antagonist. It doesn’t appear immediately. Instead the synaptic strength slowly rises over the course of some thirty minutes. Since there is no special calcium signal happening at the particular stimulated synapse it seems like maybe this form of LTP is kind of a universal cell-wide potentiation. On the other hand, it could be that there is a local calcium-independent process that marks the stimulated synapses and allows specific potentiation. Retrospectacle had a brief discussion of some of this stuff related to long-term depression last week. Reference: Raymond CR. Redman SJ. 2006. Spatial segregation of neuronal calcium signals encodes different forms of LTP in rat hippocampus. J. Physiol. 570:97-111.
Sunday, May 21, 2006
Interesting article in The Boston Globe which profiles researchers who suggest that variation in gut flora (the mix of bacteria) might be the cause of differences in body weight. Interesting fact: there are an order of magnitude more bacteria in your gut than cells in your body. Also, to my knowledge (hearing this from a microbiologist last year) this gut ecosystem hasn't really been replicated in the lab, so that kind sucks for a scientific understanding beyond description.
Update: Aetiology has much more.
The second point made by Eyre-Walker et al. in their paper "The distribution of fitness effects of new deleterious amino acid mutations in humans" is that "it will be very difficult to locate most of the genes involved in complex disease" (see my discussion of their first point here).
They come to this conclusion because, using their model, the variations that contribute the most to the variance of a disease-associated trait are rare (see Jonathan Pritchard's 2001 paper for a different way to come to a similar conclusion), and thus difficult to map, except in two cases: when the disease has no effect on fitness or the alleles invloved have been under positive selection. They clearly don't think those two cases are likely, though they do acknowledge they may number "a few". I disagree. We've all heard the story about the guy at night, searching for a lost coin under a streetlight because "that's where the light is". That's essentially what's happening here. They've derived the distribution of fitness effects of new deleterious amino acid mutations, so obviously disease alleles must be new. And deleterious. And amino acid mutations. Right? Ok, ok, it's true they mention a couple possible violations of those assumptions, but they still feel confident enough to say that "most of the genes involved in complex disease" can be described with their model. But here are a couple different theoretical paradigms to consider: the "thrify genotype" and "cryptic genetic variation". 1. The "thrify genotype" hypothesis. We've mentioned this hypothesis before, as a part of the ancestral susceptibility model for common disease. It was first formulated about type II diabetes and obesity, and goes something like this: imagine a normal distribution of a quantitative trait like the efficiency with which excess calories are converted to fat. Some people quickly convert all of their excess to fat, some less efficiently, and some not efficiently at all. In an environment of scare food resources, the first group has the clear advantage: in times of famine, they can live off the fat they stored from before. So those variants that predispose one to be on the right half of the curve (if we define the x-axis as efficiency) are favored. Now enter agriculture, which allows food from good times to be stored and eaten in rough times. The selective pressure for fat storage efficiency is somewhat relaxed, so new mutations that predispose one to be more on the left half of the curve can gain traction in the population. Finally, enter a "modern" diet of, most importantly, lots of sugar. In this environment, the people on the right half of the curve, with their "thrifty" ancestral genotype, well, get fat, while people with the new derived genotype don't as much. To extend the "thrify genotype" hypothesis to other traits, one needs a trait that was adaptive in an ancient environment but is neutral or even selected against now. One example is susceptibility to hypertension: in the expansion to colder climates, selection against ancestral alleles adapted to hotter climates have given rise to the current differential susceptibility to hypertension between populations of African and European origin. 2. Uncovering crytpic genetic variation (for a review, see here[pdf]). The premise here is that common disease is a result of the uncovering, through either genetic or environmental perturbation, of variation that wouldn't normally contribute to human health. That is, alleles influencing common disease are only conditionally deleterious or beneficial. Following the obesity example from above, operating in this paradigm would lead to the hypothesis that alleles that don't affect fat storage efficiency in normal circumstances do play a role when the environment is perturbed (i.e. in the presence of a high-sugar diet). One possible example: a mutation in the regulatory region of the gene coding for the dopamine transporter DAT1 (which has a frequency of ~70%, suggesting it may be ancestral) is associated with cocaine dependance. This variant slightly affects expression of the DAT1 gene under normal conditions (caveat: I mean normal tissue culture conditions), but in the presence of cocaine, that difference is amplified. Thus, the varation in expression is only made obvious in an altered condition. If this paradigm holds, it would mean that many of the alleles (either derived or ancetral) contributing to common disease are neutral except in certain circumstances. So if the environment that exposes this variation is our modern environment (long lifespans, high food availability, etc.), the alleles have been neutral for most of history. In general, Eyre-Walker et al. are operating in a world where an amino acid-changing mutation arises with a certain selection coefficient, and where this selection coefficient is constant until the mutation is removed from the population. If common diseases are caused by these new, deleterious amino acid changes, their conclusion that it will be very difficult to locate disease alleles is fair. But, as I've tried to show, other models for the allelic architecture of common disease are certainly plausible, and perhaps even more likely. Labels: disease
Saturday, May 20, 2006
Lordi wins Eurovision song contest...
Update from Razib: Finland vs. Sweden re: educational performance. Take home message: Finns are nerds! I know Razib was rooting for them. 
Ron Bailey has an article in Reason, Slouching toward a DNA database nation. I would like to argue that becoming a DNA database nation is not in itself a bad thing, and that there are better and worse ways to go about doing it.
First, what information is in a DNA database? From what I can tell, most DNA profiling is based on STR (short tandem repeat) sequences. There are something like 13 to 18 standard STRs that are typed. Each STR is a unique locus in the genome (like a gene) that has variant alleles; most have from 15 to 50 alleles. Genotyping this panel arguably produces a uniquely identifying signature, which can also be used to infer close kinship relationships, such as paternity. The obvious applications of such a database are identifying the source of biological (crime-scene or disaster) samples and establishing kinship (paternity). The largest such databases are related to law enforcement (the FBI), and most individuals in the database were sampled at arrest or conviction. DNA samples can be collected non-invasively, and commercial platforms exist for high-throughput genotyping. Privacy is the primary concern surrounding the existence of such databases. Civil libertarians imagine a variety of misuses for such databases. Without bothering to list them, I will say that I am sympathetic to some of these concerns. However, I think the solution is obvious: add everyone to the database. This has a number of advantages: * barring technical problems, samples would always be matchable to their source * finding an individual in the database would no longer imply that they had a criminal record * the possibility of two individuals having the same profile can be known empirically * paternity can be known for all children * the temptation for police to perform DNA dragnets will be eliminated Let me suggest further refinements: make the profiles anonymous to government authorities until a warrant is granted on the basis of judge finding that the DNA sample was acquired legally (or something like that). Also, destroy the samples after the profile is created. I am skeptical about the intermediate-term prospects of Gattaca-like fast DNA profiling for identity verification. However, it might be possible to get next-day identity verification for things like bank loans, drivers license renewals, etc.
Global Beauties got their shit together and finally put up a facebook of sorts for the contestants we know about. Despite the vagaries of image quality (some of these are glamor shots, some are only stills of the victory crowning, etc.), my 0-10 ranking below the fold (no more than 5 seconds viewing each contestant).
Update: Other opinions. Albania 7 Angonla 5 Aruba 7 Bahamas 5 Belgium 5 Bolivia 6 Brazil 6 British Virgin Islands 4 Bulgaria 4 Canada 8 Cayman Islands 5 China 7 Colmbia 7 Costa Rica 7 Croatia 5 Cyprus 8 Czech Republic 6 Domican Rep. 7 Ecuador 7 Egypt 6 El Salvador 7 Estonia 4 Ethiopia 5 Finland 6 France 7 Germany 7 Ghana 4 Greece 7 Guatemala 6 Hungary 7 India 6 Israel 7 Jamaica 6 Japan 7 Indonesia 6 Korea 6 Latvia 6 Lebanon 6 Malaysia 5 Mauritius 6 Mexico 7 New Zeland 7 Nicaragua 8 Nigeria 5 Northern Marianas 7 Norway 7 Panama 7 Paraguay 8 Peru 6 Philippines 6 Poland 8 Portugal 6 Peurto Rico 7 Russia 7 Serbia & Mont. 7 Singapore 5 Slovak Republic 7 Slovenia 7 South Africa 6 Spain 8 Sri Lanka 6 St. Vincent & the Grenadines 6 Sweden 6 Switzerland 7 Thailand 7 Trinidad & Tobago 7 Turkey 7 Turks & Caicos 4 Ukraine 6 Uruguay 6 USA 7 Venzuala 6 US Virgin Islands 6 Zambia 5 Comments: I think picture quality had an affect on who I gave an "8" to. I was tempted to give a 9 to Miss Nicaragua, but overall, the metric here is relative and I don't think she was a full point higher than the others I gave an 8 to. My own impression is that the variance here was less than in previous years, and was surprised by how few low scores I gave out. Props to China for sending an attractive candidate this time! After spending some time in the Hong Kong airport my impression is that for some reason East Asian nations don't send their "hippest and hottest." I think that the black and white photo worked against Miss Norway, otherwise I might have been bedazzled by her golden locks. Miss Finland might be an alien, but Miss Estonia is a canine. This isn't necessary from precedant, so I don't know if they weighted it toward ugly congeniality or what. India did not represent (sorry browns). In the "above its weight class" I'd give that to Northern Mariannas Islands (though I suspect that the contestant might be a black American). I guess Poland makes up for the other non-Russian Slavic groups, who did not represent (OK, I'm talking about Bulgaria).
Friday, May 19, 2006
I was going to mention this before, but I forgot. Recently I went to a talk given by a physical anthropologist, and he mentioned how humans are superior as far as large mammals go in maintaining a high level of function during the heat of the day. Offhand, he offered that if you took a 'cold adapted' Inuit and placed them in a subtropical desert they could acclimate pretty quickly. On the other hand, he suggested that the reverse was not true, that if you took a person from a tropical climate they won't adjust as well physiologically in a frigid clime. One could say this shouldn't be surprising, the hominin lineage has been predominantly a tropical one, and H. sapiens are presumably a recent arrival from Africa to much of the world. So one might accept that our heat adaptations are ancestral characters that non-tropical peoples have retained because of the vicissitudes of phylogeny. I doubt it is that simple, I suspect that there are other benefits of our heat adaptations, probably relating to cognitive function, which constrains the loss of these traits in non-tropical people. Of course the addition of adaptations to the cold as derived characters makes sense. When I asked for the source, I was told that this sort of information could be found in Human Biology. I haven't been able to get a hold of this text, but he told me that most of the references are to studies from the earlier half of the 20th century done by the US military (probably black-white comparisons?).
In any case, these findings shouldn't surprise too much, though the asymmetry was something I hadn't expected. But, it seems likely that disease was a much better trick at keeping northern peoples out of places like Africa than climate. But, this might have some bearing on why northern peoples invaded southern regions more than vice versa. Mind you, I don't think simple biological explanations are the predominant underlying component here. I've already pointed out the World Island tends to taper along many of its peninsulas in a north-south fashion so a form of geographic determinism probably plays a role. Additionally, a great deal of the conquest over the past 4,000 years has likely been in the form of steppe barbarians who descended upon civilized peoples whose martial skills were not up to par. The Central Asian steppe is in a northern clime, but the history of peoples like the Oirat Mongols or Jurched/Manchu also suggests they ventured north and displaced hunter-gathers in Siberia during some periods of history. Of course, the encroachment upon circumpolar peoples is not as notable because circumpolar peoples, like the rat-eating Finns, didn't have an indigenous literate civilization to record the onslaught from the peoples to the south. The reality is that it seems steppe peoples have periodically overwhelmed cultures to the west, east, south and north. Nomadism seems to occupy a "happy" martial medium between the hunter-gathering lifestyle and settled farming.
Yann Klimentidis points to a preprint of a paper by Eyre-Walker et al. coming out in Genetics entitled The distribution of fitness effects of new deleterious amino acid mutations in humans. The abstract finishes with this:
We argue that the relaxation of natural selection due to modern medicine and reduced variance in family size is not likely to lead to a rapid decline in genetic quality, but that it will be very difficult to locate most of the genes involved in complex disease These are two possibly interesting points; let's take a closer look at both of them: 1. Decline in fitness due to medicine (I've never heard the argument about reduced variance in family size, and it's bizarrely never mentioned again in the article). This is a classic worry of geneticists, dating back to god-knows-when but popularized as the notion of "genetic load" -- the difference in fitness between the average individual in a population and some ideal. The argument here is that some mutations that would be deleterious in an "ancient environment" no longer have a fitness cost due to medical advances, and thus are free to drift through the population. If by chance the "ancient environment" were to then re-assert itself, most people would have such a low fitness that it could be the end of the population. Of course, I sincerely doubt anyone stays up late at night worrying about this possibility. But what people occasionally stop to think about is a more subtle argument: an increased genetic load means an increased dependance on technology to offset the potential fitness costs. An example: imagine there's a genetic disease that in the past lead to decreased fitness, but is currently treatable with a little pill. If the disease becomes more common (it's not being acted on by selection, after all), more and more people will need the little pill, and, in the long run, the entire population could theoretically be affected. Thus, efficient treatment of disease could paradoxically lead to more disease and more money being spent to treat it. This is somewhat more reasonable a fear, but still unjustafiably paranoid. Most populations are already dependant on technology to one degree or another (and imagine the train wrecks our genomes are because of fire or agriculture...those innovative bastards!), and the use of technology to temper the unpredictable or inconvenient effects of the environment could probably be considered a uniting theme in all of human history. And of course, genetic engineering could (in theory, at least) render this point moot in the long run. But anyways...Eyre-Walker et al. take their estimate of the number of deleterious mutations per person per generation (1.8) and their estimate of the average effect of a deleterious mutation (a loss of fitness of 4.4%), and come up with a maximum "decline in genetic quality" due to relaxation of selective constraints of 1.8*4.4= 7.7% (compared to some ideal population). And given that complete relaxation of selective contstraints is implausible and they think they've overestimated the average effect of a deleterious mutation, they decide the "decline in genetic quality" is likely to be more on the order of a couple percent per generation. Eh. Take from that what you will. One last thing that essentially sums up why I find this whole thing a bit silly: the subheading they write this all under is "Decline in fitness". But if you define fitness in any classical manner (reproductive success, for example), medicine doesn't lead to a decline in fitness at all, but often an increase. If they wanted to be precise, the subheading would be "Hypothetical decline in fitness were medicine to disappear overnight" or "Decline in genetic quality if we consider 'quality' a measure of adaptation to an ancestral environment". When you think about it like that, it's a lot less ominous. Imagine the decline in fitness if we went back to being hunter-gatherers. Modern medicine, like agriculture, is now part of our environment (at least where I live), and our genomes will slowly adjust to that fact. Nothing to worry about there. (This has gone on a little long, so I'll address the claims about finding disease genes in a separate post)
Thursday, May 18, 2006
Saw a talk yesterday by a guy named David M. Eagleman. He’s down at UT-Screwston. He is involved in that tricky cognitive neuropsych / philosophical area that you can talk and write about for a long time and end up with a more convoluted version of the same idea you had to begin with. Apparently worked with Crick some before he passed away. This stuff isn’t exactly in my area, but it’s kind of interesting, so I will give you the rundown of what I understood.
How is it that we integrate information from all these different sensory modalities into a coherent continuous conscious experience? It’s bad enough that sound is processed in a different place in the brain than vision, but also the transmission and processing speeds for these two different modalities are different. You’d think our sensory experience would be smeared, but it’s not. In fact we can tell which event of two came first even when they are in the milli- to micro-second range. Eagleman points out that of course its very important for us to know which came first because that’s how we determine causality. His example is that when you’re walking in the woods you might have cause to be anxious if you hear a twig snap before your foot hits the ground. In fact, it appears that motor interaction with the world is one of the best ways to calibrate the system. We’re trying to pull off this incredibly important task with shoddy equipment. We simply can’t expect visual and motor information to show up at the same time. We can’t even expect events in the same sensory modality to show up at the processing centers at the same time. For instance, cells in one of the first stopover points for visual information (the lateral geniculate nucleus of the thalamus) respond at different times to different-intensity light flashes. The first response is some 80 milliseconds slower for dim flashes than for bright ones. So our integrated subjective experience has to live in the past a little bit. Eagleman calls this post-diction. Stuff from after a salient event is integrated into the perception of the earlier event. He demonstrates this just within the visual system using a temporal illusion called the flash-lag effect. You can view some movies and get his explanation here. In fact, check out his whole webpage if you’re interested in this stuff. He was a literature major in undergrad, which I guess translates to some facility with communication. Anyway. The flash-lag effect is this: When a flash of light is shone in the middle of a ring that you are watching, its perceived relation to the ring depends on where the ring is directly after the flash. It seems we if we scootch our perception of the ring movement forward in time to map onto the flash better or vice versa. Stuff from ~80 ms after the flash is projected back in perceptual time, and yes he does find it interesting that the number ‘80’ shows up here and in a number of other cognitive timing paradigms. He talked about a lot of other experiments and illusions dealing with how the system works intra-modally versus cross-modally and how intentional motor acts are good markers for temporal integration because we just did something and dammit! we expect something to happen. He even had some fMRI data from during the perception of these temporal illusions, but it was still under review, so I doubt he wants be blurting it out across the blogosphere. The basic idea is here though. We are forced to balance between our need to have the highest temporal resolution possible to get causality right and the fact that our equipment just isn’t really up to snuff. So we do the best we can with what we got and it works pretty well blurring the world in little 80 millisecond chunks until some meddling cognitive scientist comes along and mucks things up. Here’s a review from late last year for free in Journal of Neuroscience.
...with the wrong guy is available here, in case you missed it. Destined to be a classic.
Look at the number of entries in Wikipedia for Finnish porn stars vs. Swedish porn stars vs. Hungarian porn stars. This list kind of surprised me. Now I know why he's Peter 'North.'
Wednesday, May 17, 2006
In the comments of a recent post, I and a couple other people mentioned Type II, or insulin independent, diabetes. You might assume that the existence of a type II means there must be a type I and, well, you'd be right.
The difference between type I and type II is fairly straightforward: type II diabetes is characterized by a loss of sensitivity to the hormone insulin, while people with Type I diabetes don't make any insulin to begin with. The reason for this lack of insulin (in people with Type I) is, broadly speaking, that the cells that make it have been destroyed by their own immune system. The genetic basis of type I diabetes is murky, but it's known that there are risk factors in genes involved in immunity and the production of insulin, as one might expect. But there's also evidence of a strong environmental role (these points are essentially copied from the abstract of this paper): 1. a pairwise concordance of type 1 diabetes of <40% among monozygotic twins 2. a several-fold increase in the incidence over the last 50 years 3. migration studies indicating that the disease incidence has increased in population groups who have moved from a low-incidence to a high-incidence region. In this context, it's interesting that a new study looking for genetic susceptibility factors in type I diabetes fingered IFIH1, a gene coding for a protein the authors descibe as "a sensor or pathogen recognition receptor for viral infection" The implication is thought-provoking: the environmental factor (or one of the environmental factors) that "pulls the trigger" on the autoimmune destruction of insulin-producing cells may be a virus, but genetic factors in the host determine whether the response is catastrophic or not. See also: Agnostic on No Two Alike and developmental noise. Labels: disease
Brain size not linked to Microcephalin and ASPM?. Normal Variants of Microcephalin and ASPM Do Not Account for Brain Size Variability:
Related: Earlier ASPM & MCPH1 articles.
It has been a while since I asked GNXP readers to introduce themselves. If you are inclined, the comment box is yours.
Tuesday, May 16, 2006
Steve has been noting that multicultural societies are not particularly vibrant, while monocultural ones often are. I am not entirely unsympathetic to this idea, but,
1) One could quibble with defining Athens as monocultural, seeing as how metics formed a substantial portion of the population. Men like Anaxagoas and Aristotle are not trivial. But, they were Hellenes. 2) What about Christianity? Hebrew theology + Greek philosophy + Roman organization = Christianity? Right? In short, that's what we have to thank multicultural empires for: world religions (i.e., Syrian Christianity + Arab polytheism = Islam, Indian Buddhism + Daoism = Chan Buddhism, the examples are legion). Whether you think this is a good thing or bad, that's up to you....
Here is an article from today's London Times by Theodore Dalrymple, which as usual is a good enough reason for reading it.
Neven Sesardic is a philosopher of science at Lingnan University in Hong Kong. In an earlier thread I recommended his new book Making Sense of Heritability. From the dusk jacket:
There is actually nothing new here of substantive scientific interest to devoted GNXPers. Read it instead for the pleasure of watching the numbing anti-hereditarian catechism of "heritability-estimates-are-meaningless" skillfully and methodically dismantled by a sharp and incisive mind. This is an expensive academic book. You can find free copies of Sesardic's journal papers on these matters here.
Monday, May 15, 2006
Yo.. Ya’ll can’t do it like ‘Tay do it.. Can’t stop, won’t stop like a Ford Explorer without the brake fluid.. - Phonte
Around the same time as Foster and Wilson were reporting that recent event sequences in the hippocampus replay in reverse during subsequent sharp-wave ripple (SWR) events, O’Neill and colleagues were reporting that SWRs occur even as a rat is wandering around and that these seem to lead to more coordinated firing in a post-exploration sleep session. I didn’t quite explain what SWRs were before. The “ripple” part is the 140-200 Hz oscillation measured in the CA1 sub-region of the hippocampus. The sharp-wave (a self-organizing burst of activity) begins in the recurrent networks of the CA3 region from which neurons project to the CA1. The strong stimulation from the sharp-wave induces synchronous, oscillatory CA1 and CA3 firing. SWRs can happen during exploration, usually in brief pauses in theta oscillation (the predominant network activity during exploration ~10 Hz oscillation) when the rat is slowing its pace, but can also happen overlaid with theta. Rats are enjoying a leisurely stroll across campus, lost in thought. These exploratory ripples (eSWRs) are smaller amplitude, shorter, and less frequent, but with a higher oscillation frequency than ripples during immobility (iSWRs) and sleep (sSWRs). CA1 pyramidal cells are brought into tighter synchrony during SWRs than during theta oscillations, or at least more of them fire together in short time windows. Synchrony seems to indicate some sort of network level organization and communication between synched cells. They also determine that place rate-map correlation (the tendency for a place cell to fire more in one spot over others) is worse during eSWRs compared to during theta oscillation. I think the point of all this is to show that eSWRs can muck up the current place field map (potentially to change it?). That is not to say that eSWRs don’t have place-specific firing properties. In fact, the authors go to great pains to show that eSWRs increase firing rates in a place-specific fashion. The peak firing rate for a place cell inside its place field during an eSWR is actually higher than that from adding (“in field” baseline firing rate) + (eSWR peak rate out of field). I take this as saying that place cells that are “in field” have some special response to the eSWR, which perhaps identifies them for stronger integration into the current assembly. The meat of the paper is in the second to last figure (figure 7, if you have access to Neuron), which shows a ton of comparisons of correlation coefficients, which makes it very confusing to me. They are looking at shared properties of cell-pairs as a sort of proxy for network cohesiveness. Cell-pairs are grouped by correlation on one measure and then tested for correlation on another similar measure for the most part. One thing that bothers me is why on earth they would take a continuous variable like a correlation coefficient and set a seemingly arbitrary threshold for dividing cells-pairs into groups rather than just analyzing it by regression. The categorization variable is the instantaneous firing rate correlation (IFRC). This is exactly what it sounds like: How similar is the firing rate at various times between these two cells? When cells are grouped into hi-IFRC or low-IFRC groups during eSWRs and then tested for IFRC during sSWRs before (referred to as pre-sleep) or after (post-sleep) the exploration session, the first thing you notice is that IFRCs are generally weaker during sSWRs (less coherent network activity?). The second thing you notice is that the difference between hi- and lo- IFRC pairs is much bigger after exploration when the rat is sleeping in the explored environment than before the rat was ever exposed to this environment. The same thing happens if you group cell pairs by place-field similarity rather than IFRC. It seems that something is happening during exploration to arrange cells into place representing assemblies that are then reactivated in subsequent sleep. The story here seems to be that when a rat first enters a context it doesn’t really have a map for it set up. As he wanders around a little, place cells start to sort of arbitrarily pick spots. Rather than having the place cells just sort of fluctuate in and out of representing this spot or another, these SWRs come in and whack the system so hard that they alter synaptic weights. Cells that were inching towards representing the same place have an opportunity to link-up and coordinate firing a little, builiding assemblies, whereas cells that were borderline get the boot. Previously, this was presumed to happen in sleep and was supposed to coordinate hippocampal cells with neo-cortical cells in order to consolidate the spatial memory, but this paper and the previous one I covered (Foster and Wilson, 2006) show that cells can do this during waking hours. This is probably good if you went somewhere this morning and want to return in the evening, but it deprives sleep of a special role, so sorry folks we still don’t know what sleep is for. References: O’Neill J. Senior T. Csicsvari J. 2006. Place-selective firing of CA1 pyramidal cells during sharp wave/ripple network patterns in exploratory behavior. Neuron. 49:143-155. Foster DJ. Wilson MA. 2006. Reverse replay of behavioural sequences in hippocampal place cells during the awake state. Nature 440:680-683. PDFs for you: Here are PDFs of the O’Neill et al. and Foster and Wilson papers.
Just a thought. The recent data coming out of genomics, and especially the HapMap, is suggesting that there is a lot of selection in Eurasian populations over the last 10,000 years. So, on the one hand we have loci 1-40 in Chinese being selected for, and loci 20-60 in Europeans being selected form. If an appreciable number of 1-20 and 40-60 are dominant, that is, you gain function from only one copy, then European-Chinese crosses could benefit from complementation. Some of these loci will impact wholly different phenotypes, so that hybrids could hoard talents, kind of like "What if Superman & Wonderwoman mated?" But, another issue that I am wondering about is the possibility that hybrdization could increase dosage and so shift upward the median value of a common phenotype. Because the two populations have found different ways of scaling the same adaptive hill the hybrid can snatch both tricks and double their productivity. Of course, a lot of the recent selection probably has some negative side effects (think of Ashkenazi "overclocking"), so hybrids would also possibly collect a host of susceptibilities and minor ailments. I mean, look at how pensive Agnostic looks :)
Addendum: Admixture of two distinctive populations is liable to increase variance as the number of genomic permutations proliferates and loci become more polymorphic. Does that imply that variance in fitness will also increase? (addressing the issue of fitness in the comment boards should be prefaced by what you mean as fitness!)
In a thread below Michael brought up the issue that northern Europeans seem to like breasts and southern Europeans butts (ok, the thread wandered all over the place, but let's take this as the generalization). Normally, I'm skeptical of the long term persistence of cultural traits, but I once read a book on the history of sex where it mentioned that ancient Greek prostitutes padded their asses and paid homage to Aphrodite callipygia, so I'm not going to totally dismiss this.
So, a question a) anyone know of any studies on this topic in regards to what men prefer? Some of this is obviously culturally mediated, I doubt Chinese men are obsessed with feet anymore, for example. b) anyone able to tell us about the general preferences of groups like Arabs, browns, southeast Asians, etc.? c) I know that readers won't be able to help it, but please try and keep the extrapolation from personal preference to a minimum, I'm really not interested in your own likes & dislikes and how you can't imagine how anyone would differ from your rank order of traits :)
So, my computer is having issues again. It was working fine for 4 days without a battery, but then all of a sudden it shut down, and now when I try to boot it up again it blinks the red battery light in the front as if it needs a battery. Put the battery back in, and it mysteriously has 26% power, and I managed to get into the computer by ignoring the "incompatible AC adapter" warning by hitting F1. In any case, I am thinking of just getting a new battery to get this computer to work as a stop gap until I get a new one (I've got a back up lap top, which is small and old, but usable). Any idea what's going on?
I spend most of my days reading about humans and our close relations, so I always get a kick out of the crazy-ass shit that goes down in other taxa. Like this, for example:
The battle over access to mates is perhaps one of the most extreme conflicts in evolution. Male–male conflict sometimes becomes so intense that it can result in the death of all males in the population. For example, in marsupials of the genera Antechinus and Dasyurus, the diseases that appear to follow from excessive investment in mating can wipe out all males in the population after the mating season. But don't worry... This is not harmful to population persistence, however, as inseminated females produce new males. (link) Tangentially related: Sperm competition and its handiwork
In today's London Times Anjana Ahuja reports on a recent study by anthropologist Kathleen Heath on the bloodthirsty habits of the medieval English kings. (I think I've also seen this reported somewhere else, but I can't remember where.) The gist of it is that although they killed a lot of their relatives, it was consistent with their own genetic interests to do so: they never killed their own direct descendants, and their aggregate genetic relatedness to their victims was always much less than that of their direct descendants. So if by killing collateral relatives they protected themselves and their direct descendants they were promoting their own inclusive fitness.
Well, maybe. The problem is that genetic relatedness 'decays' so quickly with distance that you would have to kill an awful lot of distant relatives - e.g. five nephews or nine first-cousins - to outweigh the benefit of saving yourself or two offspring. So a monarch would have to be very bloodthirsty (or paranoid) indeed to fail the test. Interestingly, Heath suggests that the Ottoman Empire would show a similar result. If it does, I would be more impressed. If I recall correctly, the first act of every new Sultan was to have all his brothers strangled with a bowstring. These would usually be only half-brothers, so it would be necessary to kill at least five of them to break Hamilton's Rule, but on the other hand, there were so many concubines in the Seraglio that there might well be sufficient potential victims to put the hypothesis to a real test. [Added: having just written this, it strikes me that Sultans would also potentially have a very large number of offspring, so they would have to kill an even larger number of brothers to fail Heath's test.] [Added #2: but then again, if your brother becomes Sultan he also potentially has a large number of offspring, so his genetic value to you is proportionately greater. It all gets quite intricate...] [Added #3: the other report that I vaguely recalled was at Steve Sailer's blog. The study was also discussed at Razib's Science Blog. Returning to the point in Added #2, I think the (approximately) correct analysis is as follows. Suppose that a non-Sultan has on average A surviving offspring (where A is usually about 2). Suppose also that a Sultan who eliminates his rivals has A + K surviving offspring, where K is a substantial but not huge number (say around 20). Suppose finally that a Sultan has N half-brothers, all of whom have an equal chance 1/N of displacing him if he does not kill them first. The genetic value of a new Sultan to himself can be expressed as 1 x (A + K). The average genetic value to him of each of his half-brothers, if they are allowed to survive, can be expressed as 1/4 x [A + K/N]. The point of 'K/N' is that a brother who takes over as Sultan will have an extra K offspring, but each brother has only a 1/N chance of doing so. Given these assumptions, it will pay a new Sultan to kill his brothers unless N>X, where X = 4[(A + K)/(A + K/N). However, this is still only a superficial analysis. The Sultan would really need to know whether or not the custom of killing brothers will continue, as this affects the value to him of all his brothers. It is possible that there are two different Evolutionarily Stable Strategies: maybe if the custom of killing brothers is expected to continue, it pays the Sultan to kill his brothers no matter how many there are, whereas if the custom were likely to be abolished, it would pay him to let them live.]
Sunday, May 14, 2006
Sometimes a picture really is worth a thousand words. Other times you need a thousand words to understand the picture.
In following a trackback I came across a translation of the Chinese text:
 Updated">
Just talked to an adoptee from Korea who was force-fed milk as a child because her mother wanted her to overcome her lactose intolerance.
Saturday, May 13, 2006
From a vicious profile in The Times:
"We men are like the Cro-Magnon people of prehistoric times," he once told me. "We're always hunting and wenching. But, at the end of the day, we always go back to our caves. For my part I need this cave to feel at ease with myself. Without it I would be as unhappy as could be." 1. "Wenching"? Did somebody use an online translator here? 2. Is it 2007 yet? (Via ALDaily)
Friday, May 12, 2006
Miss Finland talks like an alien baby!!! (oh wait, that's just suomen kieli)
posted by
Razib @ 5/12/2006 11:31:00 PM
Watch Miss Finland, Ninni Laaksonen, talk like a freaky alienoid infant. Here are some pics of her non-earthling self. Oh, and the new Miss Sweden looks like a depigmented Jessica Alba (who is 1/4 Danish) to me.
Update: It has been brought to my attention that 1) Miss Finland might be of Swedish ethnic origin (apparently the interviewer asks Miss Finland if it is an advantage to be a Swedish Finn in the competition), and Miss Sweden is probably a 1/4 each: Finnish & Russian.
A comment on another weblog:
On the first point, well, not really. On the second point, what jargon is confusing??? I'll be away from the computer this weekend, but I'm curious as to what comments about confusing jargon are offered in the discussion thread....
Dienekes points me to this piece in The American Journal of Physical Anthropology which concludes that there isn't greater sexual dimorphism in skin color in areas where social/sexual selection could presumably operate because of relaxation of natural selection. Remember that sexual dimorphism tends to evolve very slowly (you need to have sex-linked or developmentally modified loci since men and women share almost all the same genes [except for the Y]), perhaps an order of magnitude more slowly than standard phenotypic evolution given the same selection pressure.
Labels: Pigmentation, sex differences
Thursday, May 11, 2006
I put my thang down, flip it, and reverse it. – Missy Elliot
I have been invited on as an author at Gene Expression, so this post will go up at The Genius and at GNXP. To GNXPers: Hello. I'm a grad student interested broadly in neuroscience and genetics. Hopefully I'll be adding signal and not noise. This post is about an electrophysiological phenomenon in the hippocampus that may play a role in locking in lasting memories of event sequences. Sharp-wave ripples in memory consolidation are sooo hot right now. I have been following the leads in this Wendy Suzuki mini-review to some really interesting multi-cellular recording studies in the hippocampus. She focuses on three recent papers, one of which I discussed here but hope to revisit. The most straightforward paper of the three is this report from Foster and Wilson. They show that cells in the hippocampus replay firing patterns that represent the places a rat has just been, but they do it in reverse. Another good avenue into the paper is this commentary by Colgin and Moser. To get the paper you have to get a grip on sharp-wave ripples and place cells. To get the theoretical issues you probably need to know more about dopamine and its role in reward learning. I’ve discussed place cells at some length here, here, and here, but for the uninitiated here are the basics. Place cells are found in the hippocampus and closely related areas (such as the part of the cortex that feeds directly into the hippocampus). They have place fields, just like cells in the visual processing system have receptive fields. They fire more as the rat moves toward the spatial location that is at the center of the place field. There are enough place cells with different place fields to make up a full-coverage map of a rat’s immediate environment. When the rat is moved to a new environment a new configuration of place cells comes online and represents a map of that environment. Observations in this vein led to a theory of the hippocampus as a cognitive mapping system espoused by John O’Keefe and Lynn Nadel in a book called The Hippocampus as a Cognitive Map in 1978, which I was amazed to find that you can download the full-text of for free here and it’s only 3.3 MB. Sharp-wave ripples (SWRs), I am only beginning to become friends with, but they are super-interesting. So when you are recording in the extracellular space in most layers of the hippocampus there are network-level potential sweeps that occur in different behavioral states. An example that I’ve discussed before is theta oscillation, in which the extracellular potential fluctuates at a rate of ~6-12 Hz or an oscillation about every 100 milliseconds. Theta oscillations usually happen while a rat is walking around and exploring. SWRs are measured the same way, but they are much higher frequency and usually happen at times when theta is not happening, for instance during consummatory behaviors (chowing down), waking immobility, and during slow-wave sleep. Some of the most compelling research with regard to SWRs has shown that patterns of place cell activity that occured during the day are recapitulated in miniature during SWRs in slow-wave sleep, and that these SWRs are coordinated with another electrophysiological phenomenon called spindles in the neocortex. This latter finding is very interesting in terms of the standard systems consolidation view of memory, wherein memories intially stored in the hippocampus are moved over time to permanent cortical storage sites. Foster and Wilson are not content with replay during sleep. They recorded from pyramidal cells in the CA1 region of the hippocampus using electrode arrays of 17-18 tetrodes (I'm not sure if you need one tetrode for each cell of if they get 72 cells out of this deal) as rats ran back and forth on a linear track to retrieve goodies at either end. First they show a set of place cells that represent the whole journey (i.e. one fires in Los Angeles, the next in Las Vegas, the next in Flagstaff (yes, we're going to Amarillo)). But once the rat gets to Amarillo and chills out, the all the cells spike together...well not exactly together. They fire at a greatly compressed timescale in reverse order with regard to the recent journey. It's like the rat is thinking back over it's trip starting with the most recent event. Several of these reverse replay events can happen per resting period, and they happen during SWRs. So somehow this fast wave of activity sweeping across the hippocampus must be working its way backwards from the most recently spiking cell assembly to the most remote. Now that I'm thinking about it, I wonder if the spikes are setting up some sort of slow-decaying process that makes a neuron more reactive, like a release of intracellular calcium stores or maybe a signal transduction event leads to increase of current flow through ion channels or something. Then the cells most recently activated would be closest to threshold when the SWR activity shoots through. The authors actually propose something quite close to the above now that I look back at their model. Although they don't bother with a fancy mechanism, they just assume that recently activated cells are at a higher level of excitation. The reason why I skipped over that simpler idea is because it's my impression that cells that have just fired are hyper-polarized (refractory period) and less likely to fire, so I needed a process that didn't just rely on action potential mechanics. It's probably best to assume that they took this into account since they are smarty-pantses. We get some intriguing theoretical stuff when the authors attempt to explain why this reverse replay would be useful. It goes something like this:
There are some good reasons to believe in this model. The authors note that other (non-dopaminergic) parts of reward circuitry can fire in synch with SWRs, and we know that in some cases the presence of dopamine determines whether or not activity-induced changes in the neural circuitry actually stick. The only problem I have is that I have a hunch that these reverse replay events might happen even if there wasn't a reward. The authors report that several of these SWRs can happen per stopping period, and the rat isn't getting a new treat for each one. Sooo why are they happening in the absence of reward? Maybe you can just explain the phenomenon in terms of cellular excitability in reverse order, and you don't need to tack on the reward issue to explain the phenomenon but it does add an intriguing twist. Also, I was thinking about the idea that these are in reverse and the ones during sleep are forward. I haven't read the papers about the sleepy-time ones, but I wonder if its worth looking for a slow reverse replay prior to the SWR-forward play. Seems like you might find something like that if the reason SWRs activate cells in a particular order is because of their recent spiking history. References: Suzuki WA. 2006. Encoding new episodes and making them stick. Neuron 50:19-21. Foster DJ. Wilson MA. 2006. Reverse replay of behavioural sequences in hippocampal place cells during the awake state. Nature 440:680-683. O'Keefe J. Nadel L. 1978. The hippocampus as a cognitive map. Clarendon, London.
Three papers in the January issue of Nature genetics, "Common deletions and SNPs are in linkage disequilibrium in the human genome", "Common deletion polymorphisms in the human genome" and "A high-resolution survey of deletion polymorphism in the human genome" each examine common structural variation in the human genome.
A number of papers have previously reported that intermediate-scale "structural" (e.g. insertions, deletions, rearrangements) variations "are prevalent enough to be an important source of genetic variation between individuals". One question was whether these intermediate-sized variations occured once during human history and shared by descent (like most SNPs) or whether they are like a class of recurring mutations that delete, insert or rearrange large portions of the genome and subseqently cause disease. The Perlegen paper gives the best summary: common deletions and SNPs ascertained with similar criteria have essentially the same distribution of linkage disequilibrium with surrounding SNPs, indicating that these polymorphisms may share evolutionary history and that most deletion polymorphisms are effectively assayed by proxy in SNP-based association studies.
The BBC reports on a new study of Neolithic injuries:
A survey of British skulls from the early part of the New Stone Age, or Neolithic, shows societies then were more violent than was supposed. Blunt instruments such as clubs were responsible for most of the trauma. If this can be seen as news is doubtful, as the subject has already been explored extensively in a long-running and widely viewed documentary series... 
This week's Nature has an excellent news article on epigenetics.
We all know humans share some large and horribly impressive percentage of our genome. But we're all different, and studies (using resources like the HapMap) of those under-appreciated percentages of the genome we don't all share should give us a clue to why that is. But identical twins share an even more impressive percentage of their genome. How, given the fact that they're, well, identical, can they be different? The news article cites this study and describes it: By studying 80 pairs of identical twins, ranging in age between 3 and 74, Esteller's team found that epigenetic differences were hardly detectable in the youngest twins, but increased markedly with age. These changes had a striking effect on gene activity: the number of genes that differ in activity between 50-year-old twins was more than three times that in pairs aged 3. "So we are more than our genes," says Esteller. "Not only is the DNA sequence important but also how gene activity is regulated in response to environment. This might explain why many identical twins have different susceptibility to disease." And the promise for better understanding of phenotypes is there: [Epigenetics] should also fill some big gaps in our grasp of how the environment affects a creature's constitution — epigenetic changes explain how simply altering the diet of a pregnant mouse, for example, can completely change the coat colour of her pups, or even alter their response to stress. As of now, it's technically difficult to get a genome-wide view of methylation, but arrays to do just that should be just around the corner. See also: Microarray-based DNA methylation profiling: technology and applications
A new paper in PNAS shows how the power of cheap(er) sequencing is changing how a lot of people approach questions.
Here, the question is this: once a person is infected with a bacteria, obviously the bacteria population doesn't stay static; your body and whatever you put into it are a different environment, and the bacteria are likely to be under selective pressures. But by what mechanisms does the bacteria evolve over the course of an infection? To answer this, researchers isolated P. aeruginosa, an opportinistic bacterial pathogen, from a cystic fibrosis patient at various points over the course of 8 years. Then, they took the first and last isolates and-- hell, why not-- just sequenced them. Your average whole genome shotgun. Their results are convincing-- you're not an easy environment to live in. There was selection all over the genome, particularly against virulence factors and for drug resistance, as you might expect. They then looked at some of the mutated genes in other patients (apparently full genome sequencing is still a little too expensive to be completely routine) and found, in those genes, 5 synonymous mutations and 103 nonsynonymous mutations. That gives a ratio of a hell of a lot (keep in mind as well that there are more synonymous than nonsynonymous sites). However, outside of those few genes selected in most other patients, the genes mutated in the original patient were rarely mutated in the others. There could be any number of explanations for this, and the authors don't really speculate. However, in my mind the most chilling (or reassuring) possibility is that, after 8 years, a bacteria isn't adapted to just any human body, it's adapted to you.
I've got a dell 5100 laptop, and I've had it for 3 years now. I'm sure it's on its last legs, and I'm looking around. Anyway, I had this weird problem, and I just want to know what's going on. Over the past few days I've had issues where the computer wouldn't recognize my AC adapter. But, I noticed something weird, my battery, which has been draining and charging erratically for nearly a year now seems to often LOSE power when the AC adapters is in. For a while I couldn't even boot the computer up, the green lights would come on, and then disappear (or I'd get an error). The battery was down to zero, but the AC adapter was plugged in. Now I have the battery removed, and it seems to work OK. What's going on here? Does the computer automatically try and use the battery to jump start the system when there is a battery around? (and since my battery was at zero it just shut down?)
How reliable are empirical genomic scans for selective sweeps?
posted by
the @ 5/11/2006 01:06:00 AM
Kosuke M. Teshima, Graham Coop and Molly Przeworski from the Department of Human Genetics at University of Chicago, home to both the Pritchard and Lahn labs, present an analysis of the effectiveness of empirical scans for selective sweeps, such as those from the Moysiz and Pritchard labs.
Abstract: The beneficial substitution of an allele shapes patterns of genetic variation at linked sites. Thus, in principle, adaptations can be mapped by looking for the signature of directional selection in polymorphism data. In practice, such efforts are hampered by the need for an accurate characterization of the demographic history of the species and of the effects of positive selection. In an attempt to circumvent these difficulties, researchers are increasingly taking a purely empirical approach, in which a large number of genomic regions are ordered by summaries of the polymorphism data, and loci with extreme values are considered to be likely targets of positive selection. We evaluated the reliability of the "empirical" approach, focusing on applications to human data and to maize. To do so, we considered a coalescent model of directional selection in a sensible demographic setting, allowing for selection on standing variation as well as on a new mutation. Our simulations suggest that while empirical approaches will identify several interesting candidates, they will also miss many—in some cases, most—loci of interest. The extent of the trade-off depends on the mode of positive selection and the demographic history of the population. Specifically, the false-positive rate is higher when directional selection involves a recessive rather than a co-dominant allele, when it acts on a previously neutral rather than a new allele, and when the population has experienced a population bottleneck rather than maintained a constant size. One implication of these results is that, insofar as attributes of the beneficial mutation (e.g., the dominance coefficient) affect the power to detect targets of selection, genomic scans will yield an unrepresentative subset of loci that contribute to adaptations. They spell out the implications of their findings pretty clearly at the end of the paper: The ability to detect recent directional selection from polymorphism data depends on the recombination environment of the selected site (Supplemental Fig. S5), the dominance coefficient of the favorable allele (Fig. 5), the selection coefficient of the favorable allele (Supplemental Fig. S4), and whether the allele was favored from introduction or not (Figs. 3 and 5). Thus, if a candidate region does not stand out in empirical comparisons, it may be that there is little power to detect the mode of selection acting on it. This possibility has important implications for the interpretation of genomic scans of polymorphism data. Ultimately, we would like to use the results of genome scans to make inferences about which phenotypes were recently selected, and how selective pressures differ between populations. But we know that phenotypes differ in their genetic architectures, and thus the power to detect selection on different phenotypes may vary considerably. This raises the possibility that biological processes (e.g., GO categories) picked up in genomic scans are not those on which there was the most selection but those on which selection tended to act on new, co-dominant mutations in regions of low recombination.
Wednesday, May 10, 2006
The Finns and Swedes are arguing eye color. So, Eye colour: portals into pigmentation genes and ancestry:
One major locus, but lots of little modifier loci which an account for shades of green, gray, etc. (not to mention epigenetically derived variation). Also, remember this isn't MC1R, so that is why eye color and hair color are inherited independently. Take that Jaakkeli! You lemur-eyed freak! Labels: Genetics, Pigmentation
A few other notes from the conference:
1. Genetics of taste: Taste-blindness to PTC or PROP (bitter tasting compounds to the rest of you) is present as a recessive trait in about 30% of European-origin populations. A presentation by Beverly Tepper showed that "non-tasters" are less sensitive to a wide variety of other tastes-- spicy food, whole milk (as a proxy for the taste of fat), and red wine, to name a few. And further, "non-taster" children consumed more calories per day than their "taster" counterparts. She then tried to relate this to body weight, with mixed results. 2. Dysbindin and intelligence: another group has also apparently been studying the relationship between the dysbindin gene and schizophrenia/cognitive traits (this gene has been previously associated with cognitive ability). They looked at some measures of verbal learning and memory and found...nada. The study's not perfectly comprable to the previous one, but it's certainly not encouraging. 3. Perennial GNXP favorite Bruce Lahn...didn't show. Some British dude gave a summary of the evidence for selection on ASPM and MCPH1 instead. 4. Dutch cops. I was stopped and threatened with arrest for...crossing the street (on foot) on a red light. I seriously had hold myself back from laughing at the poor guy. Was this a joke (I've heard people from Amsterdam have a strange sense of humor)? Addendum from Razib: We've talked about taste before. There is interpopulational variance in regards to taste vs. non-taste status.
Eco de Geus gave a talk on his search for endophenotypes for IQ. A bit of background: when searching for genetic factors involved in a phenotypic trait, it's easiest to look directly at the trait itself-- that is, if the trait is a disease, grab some sick people and some healthy people and compare the two groups.
A more powerful approach, however, is to look at something closer to "gene action", as they say; look at BMI instead of diabetes or QT interval as opposed to heart disease. It's easy to imagine why this works: instead of diluting your cases with people who have disease for multiple, distinct causes, you're looking only at one of them. IQ, de Geus claimed, is a complex phenotype that would be amenable to this kind of approach. So he's been looking at various measures to bring IQ down to a more physiological level (using twin studies). But, well, he ain't there yet. Here are some results: 1. Speed of signal transmission in neurons: heritable, but not correlated to IQ 2. Processing speed (as measured by one of the waves in an EEG-- this is obviously not something I know much about): heritable, but not correlated to IQ. 3. Cortical connectivity: heritable, but not correlated to IQ. 4. Brain volume: heritable and correlated to IQ. But he said he didn't like this as an endophenotype because he found "high IQ --> big brain" to be a more plausible explanation for the correlation than "big brain --> high IQ". 5. The one test that showed a little promise was a psychological test of visual inspection time, as measured, if I remember correctly, by the Stroop test. But this was conterintuitive-- the (weak) correlation was negative (those who reacted fastest had lower IQ). So what to make of this? I, for one, know jack about this area, so don't look at me. Labels: IQ  I recently returned from the European Human Genetics Meeting in Amsterdam, and plan on writing a few posts on some of the presentations that might be of interest to your average GNXP reader. So here goes: I recently returned from the European Human Genetics Meeting in Amsterdam, and plan on writing a few posts on some of the presentations that might be of interest to your average GNXP reader. So here goes:With the completion of the sequencing of a human genome, much attention has turned to sorting out what the hell all of that DNA stuff actually does. Given that no one really knew how to figure that out, new technologies were needed. The ENCODE Project, which is currently ongoing, was designed to address that need. In the first stage, 1% of the genome was selected (some regions at random and some for specific reasons) as a sort of "testing ground" on which a huge number of groups are testing their computational and experimental methods for genome annotation. Once that 1% is annotated to everyone's content, the best methods will be scaled up to annotated the whole genome. At the conference, a speaker from the ENCODE Genes and Transcripts Analysis Group gave a talk on what they're seeing so far. Here are a couple of his most attention-getting points: 1. In the ENCODE regions, about 90% of the sequence is transcribed (from one strand or the other). For comparison, the percentage of the human genome that codes for proteins is about 2 or 3%. 2. Most genes have alternate site where transcription can start, and of these, 30% are more that 100kb away from the actual gene. In many cases, this leads to the mixing and matching of exons from different genes. I find those numbers absolutely mind-blowing (ok, ok, given what we see in yeast this shouldn't be that surprising. And I imagine some molecular biologist has known this for years, but this was a human genetics meeting-- we're a bit behind the curve). The "one gene - one enzyme" model is long gone, but it's still provides the general framework for how your average biologist thinks. In the coming years, however, there will be a sea change in how we understand the genome, The simple notion of discrete "genes" bounded by "regulatory regions" has been useful, but it's always been just a simplification; it's time for a more realistic null hypothesis.
I asked a Finnish weblog to use a language I could understand, and a reader responded with an obscenity....
Tuesday, May 09, 2006
At my other weblog I just posted something about the possibility for fitness benefits for those who are heterozygotes for the GJB2 locus. For one particular allele GJB2 causes deafness when homozygous. About 1 out of 35 southern Europeans, 1 out of 80 northern Europeans, and a large number of Ashkenazi Jews are carriers, that is, phenotypically wild type heterozygotes. Non-European populations seem to lack this allele. The south-north gradient seems to hint at long term density dependent effects, but a recent presentation at a European genetics conference suggests that carriers might heal their wounds faster! In other words, carriers might be somewhat more fit than either homozygotes, obviously more fit than the deaf, but perhaps more fit than wild type homozygotes. This is called overdominance, and the classic situation is sickle-cell anemia, where the heterozygote is protected to a great extent against malaria. It doesn't matter what the GJB2 mutation confers, but the pieces are coming together. I think many "disease" like eczema and cystic fibrosis are important because as Greg points out the frequency of the alleles which cause these problems are just too high to be attributed to mutation-selection balance (my understanding is that the rule-of-thumb for mutation rate at a Mendelian locus is on the order of 10-8, and if the a lethal recessive's equilibrium frequency is the square root of the mutation rate, we're talking 1 out of 10,000, not 1 out of 100 as carriers).
This stuff is important because if Moysiz and Pritchard papers are correct, that there is a lot of recent human selection, a subset of these are going to be overdominant alleles which haven't had time to acquire modifier loci which mitigate their downsides (whether as heterozygotes or as homozygotes). So, Mendelian diseases are important because people get sick, but they might also be a nice sketch outline of the map of recent human selection....
The Grand Slam Ranking for the Global Beauties site takes the 4 international beauty pageants and generates a composite "score." I've dismissed faux precision before, but admitted that it can be quite fun. I was going to generate some excel charts for you (e.g., plot population vs. score), but I really don't have time so I'll leave it to readers with more minutes to burn than I. Also, as expected, the Harvard of beauty pageants, Venezuela, is #1. Just to generate some controversy, I've always noticed a tendency for China to not represent, considering that there are a 500 million Chinese females [and counting down thanks to infanticide, abortion and neglect]. I also think some of you might find the links to regional/national pageants interesting. It is amusing to me that the world's only state where Hinduism is the official religion, and the constitution states that the monarch must of "Aryan" heritage, puts forward contestants than look so "Asian". While some of these women look conventially north Indian, while as expected contestants from Tibetan ethnicities look Tibetan, some with north Indian names look mostly East Asian. And of course you have the "mixed" look which seems typical from what I have seen.
Monday, May 08, 2006
I was reading Steve's blog, and he posted an email from a reader who "asserts" a lot of things. This caught my eye:
Caveat, I don't know much about foreign policy. Back a few years ago I watched the whole "warblogger" phenomenon. I don't know if I considered myself one...I was never that interested in foreign policy and didn't have strong political preferences. If you look through the archives you will see that I said things like "I am 55% in support of the war," etc. I was just going along, and occasionally I would repeat talking points I'd seen around the web to anti-warish friends. I figured that other people knew more than I did, since of course they read the news and kept up, you know, the AP, Reuters, The Jerusalem Post, The New York Times. Read those and you can comment intelligently, right? No. People didn't know jack shit. This doesn't speak to the details of the foreign policy...a lot of that depends on what your a priori values are. But, I will assert that people didn't know jack shit, because I don't think they knew non-trivially more than I did, and I didn't know jack shit. Today, I still don't. And I don't plan on knowing much more, my puny brain is focused on science, history and other things that might interest me. Then, a few weeks ago I saw a post on ParaPundit about plans to do a divide & conquer on Iran by encouraging the Azeris to rebel against their Iranian overlords. At this point, I did a Cochran and exploded with rage, because I do know a tiny bit about the history of Iran (I'm not saying tiny ironically, I don't know that much, I'm very much a dabbler) and the Azeris' Iranian overlords are...Azeris!!!! There is a reason that Azeris, a Turkic speaking people who inhabit the northwest of the modern Iranian state, won't rebel against the Iranian state, and that's because they created Iran!!! Yeah, you read that right, they created Iran! And to some extent, they still run Iran. In The Sons of the Conquerors Hugh Pope notes that a) The Iranian military is disproportionately Azeri b) The current Grand Ayatollah of Iran, Ali Khamenei, is from an Azeri family in eastern Iran (people move you know, even in non-Western countries) Some history. In the early 16th century Shah Ismail conquered Iran. He instigated the process of converting Iran to a Twelver Shia state, eradicating Sunnism from most of his domains (in modern Iran Kurds and Balouch are the main Sunni groups, Farsi speakers, Arabs, the Azeris and other Turkic groups are Shia). He was a Turk, what might be called an Azeri Turk. Until the ascendency of Reza Pahlavi in the early 20th century Iran had not had an ethnically Persian dynasty for nearly 1,000 years. Iran's ruling caste was Turkic. Now, we shouldn't perceive ethnicity as we do in the post-French Revolutionary sense. The boundaries between Turk and Persian was likely fluid, and Farsi was the language of culture. Though self-conscious Persians did not rule the historic Persia for a thousand years, Persian culture flowered. In fact, many of the early Ottoman Sultans preferred Persian to Turkish, just as Frederick the Great preferred French to German. Here's some other facts. Around 1/4 of the Iranian population is Azeri. 1/4 X 70 million ~ 17.5 million Azeris in Iran. There are about 9 million people in Azerbaijan. The leaders of Azerbaijan would be entering into political suicide if they were playing a game of Greater Azerbaijan, the secular Russian-influenced Azeris of Azerbaijan would be heavily outnumbered by religious Azeris of Iran. Today Azerbaijan might look to Turkey as its "natural" ally because of shared Turkic heritage (the division of various Turkish languages is somewhat artificial from what I gather), but traditionally the great wars between Iran and the Ottoman Empire were wars between Turks, Shia and Sunni, not Persians and Turks. Anyway, enough from me. I doubt I'll be making many political posts in the near future...but anyone saying stupid shit about stuff like "let's foment Azeri separatism in Iran" will be met with an immediate ban. This isn't quantum mechanics, all you need to do is go to Wiki or read a book. Pretend I'm Jaakkeli and Iran is Finland, and you'll get the idea of how I feel about people saying grossly and unself-consciously stupid shit. Updated: OK, Greg emailed Steve and pointed out some of the same points as above. I don't know much about foreign policy, but talking about Azeris rebelling against the Iranian state because they are resentful about Persian domination is so wrong that it makes you skeptical about either the real knowledge background that someone brings to the table or their good faith. Addendum: Anyone can feel free to correct me if they know from personal experience that Azeris resent their prominent role as soldiers and leaders in the Iranian republic and would prefer to be even bigger fish in a smaller pond.
Check out Yann C. Klimentidis weblog. Yann's research at UNM is in human evolutionary ecology and genetics.
Sunday, May 07, 2006
Battle of the beauties below the fold....
Swedish...   ...and Finnish ...and Finnish
Saturday, May 06, 2006
This is probably the most thorough coverage of the development that there are now government sanctioned women preachers in Morocco. Granted, they are not imams, but I think that development is inevitable now. One thing to note is that many smaller and Lefty Roman Catholic denominations in the USA depend on female parish employees for their day-to-day activities, but because of the special role of priests in Roman Catholicism, and its central organization, the elevation of women to roles equivalent to what has occurred in some Protestant denominations seems to be unlikely in the near future. But I think in Islam the imam is more like a rabbi or Protestant pastor, and, the lack of central authorities in the Roman Catholic mode makes the possibility of female imams more plausible. This development in Morocco is important because Morocco is a predominantly Arab country. While the rise of female imams in China is interesting in illustrating the elasticity of interpretations of Islam, when changes come to the Arab Muslim nations than you have something that reflects possible transitions toward the mainstream. It is not irrelevant the king of Morocco claims direct descent from Muhammad (though, most people in the Muslim world can probably trace their descent from Muhammad somehow if they were omniscient, and perhaps most people in western Eurasia).
Related: Here is a timeline when American Protestant denominations began to allow women into ministerships. Note that this trend is not limited to liberal groups, the Church of the Nazarene had a woman as a minister before 1900. John Ashcroft pointed this out to Patrick Leahy years ago when the latter was implicitly criticizing Ashcroft's religiouis extremism. Ashcroft contrasted his church's more open attitude toward women in positions of religious leadership than Leahy's own Roman Catholic church, and Leahy was not pleased. In any case, the point is that women won't necessarily make a church less fundamentalist, in fact, polls tend to show that women are more Creationist then men. Rather, I suspect women are more like to make the interpretation of the religion less violent. I would say pussyish, but I don't want to make this into a "unsafe space." Addendum: A celibate priesthood might also make a religion less violent. Just a thought. The Roman Catholic church's turning away from a muscular form of Christianity might track the influx of, uhm, less masculine men into the priesthood.
Dienekes points to this interesting article about the settlement of Russia by Slavs:
This is unpublished data, though it generally confirms other studies I have seen here and there. One issue though is of the idea promoted here that the settlement of the north and east of European Russia was mostly male mediated as Slavic women remained in the territories ot the south and west. There seems to be an implication here that Slavic males engaged in a mass migration, similar perhaps what we saw in Latin America with Iberian males. But, I am not clear if the granularity of the study here could distinguish this from a more prosaic wave of advance in the context of operational matrilocality. For example, if the geographic distance of the samples collected showed a discontinuity than the mtDNA would likely track that even the allele frequencies graded into each other at a constant rate. The dilution of the original genetic profile in a wave of advance scenario could also explain the "Finno-Ugric" character of extreme northern Russian profiles.
Friday, May 05, 2006
The New York Times has an interesting piece on psychometricians and the demand for them because No Child Left Behind. The only issue, which isn't mentioned, is how are scientists who know children are going to be left behind keeping a straight face designing and analyzing these tests?
A few days ago I asked why someone called me "zeeb." The reason I ask is this: "zeeb" is a common truncation of my name...the other common one is "raz." But "zeeb" outnumbers "raz" by about 25:1. Additionally, "zeeb" is the default preference of many people who I know who don't know each other, both on the web and in my regular life, so it isn't pure memetics. Additionally, as for my name, "Razib," it is actually Rajib, the Bengali form of the name Rajeev/Rajiv. It became Razib because of issues with regards to pronounciation with my kindergarten teacher. I thought this was an anomaly until I got on to the web ran into other people with the name Razib, most of them from Bangladesh (check Friendster for what I mean).
I don't know what to make of all of this. But I thought I'd make my curiosity on this issue clear.
RPM has a post about MHC and chimpanzees.
Thursday, May 04, 2006
I don't read many blogs, but I suspect some of you do. What do you read? And why?
P.S. I'm not fishing for compliments, don't offer GNXP :) Update: agnostic is asking about other types of sites.  Scientists and advocates of the National Institutes of Health (NIH) are anxious about the funding outlook for biomedical research. After the agency's budget rose from $13.6 billion in 1998 to $27.3 billion in 2003, there began what some call the 'undoubling' of its budget. The squeeze is starting to be felt. The competition for grants has become brutal, with fewer than one in 20 applications winning approval. This year the number of new grants funded by the NIH is projected to drop by almost 15 percent from the number funded three years ago.
I saw this at Aziz's blog and I signed up. It is something of an empty gesture for me because I rarely get into the personal disputes with other bloggers, though this was more semi-common in the first few years of this weblog's existence. But who knows, if there was more tension about the Wright-Fisher controversies perhaps I'd be getting down & dirty too. But I'm all for people behaving less like animals, so I signed up (OK, I mean emotional animals, something sedate like a ground sloth is tolerable because it is ignorable).
I posted some musings on this subject a few months ago (here), so I was pleased to see a sensible article by the admirable Camilla Cavendish in today's London Times. Free online version here. (For the benefit of non-UK readers, 'Boots' and 'Superdrug' are chains of pharmacies in the UK.)
Cognition and the sex chromosomes: studies in Turner syndrome
posted by
Razib @ 5/04/2006 01:29:00 AM
Cognition and the sex chromosomes: studies in Turner syndrome:
Tuesday, May 02, 2006
Your opinion solicited.
A discussion about peer review at my other weblog.
Update: Brown gaucho has a lot more commentary. Update II: Janet has more.
Monday, May 01, 2006
There's a recent paper in Nature (here), in which the authors claim that starlings show evidence of understanding recursion, the putative "core mechanism" of human language on Chomsky, Hauser, and Fitch's account (by no means consensus). I don't have university access, but there are plenty of good reviews: lay-directed ones at LiveScience and John Hawks, and somewhat technical ones at Tensor and LanguageLog.
Recursion in syntax is when a category X can be composed of another representative of the same category X, plus perhaps other stuff. For example: the rule "S --> S and S" says that a sentence (S) may-consist-of some sentence, the word "and," and another sentence, in that order. This result can feed into another application of the rule, like so: "I study linguistics and Razib studies biochem," "I study linguistics and Razib studies biochem and Darth studies psychometrics," ad infinitum, making sentences out of sentences. This is what allows human language to be so flexible -- in Wilhelm von Humboldt's classic phrase, it allows the "infinite use of finite means." [1] The links above give good overviews, so I won't waste anymore space rehearsing the details (thus, minor "outside" reading required). The take-home message is that while not a negative result for other species possessing a putatively human-only syntactic mechanism, they're not necessarily strong positive evidence either, as the Tensor & LanguageLog entries show. For all we know, the starlings used short-term or working memory, not part of a language module. Unfortunately, the three fields of cognitive sciences / linguistics, psychometrics, and evolutionary biology don't talk an awful lot to each other (though see commentaries on LanguageLog here and here for mention of potential role of general intelligence in the starling study). Hence, the possibility that the results could reflect the starlings bringing some avine version of g to bear on the task wasn't fully explored. My personal take on these sorts of studies, where the animals only succeed after tens of thousands of training trials, say more about animal psychometrics -- using general intelligence to consciously figure out murky patterns -- than about animal instincts -- effortlessly acquiring the skills & knowledge necessary to survive & reproduce. The fact that 9 of the 11 starlings learned the grammars suggests that they may be a bird version of a pretty easy IQ test. If, after tens of thousands of trials, a group of humans were trained to jump & remain in the air for several seconds of hang-time, that would say more about general athletic ability than about a basic human instinct for quasi-flight. [1] Math / Comp Sci nerds can see how this works in old posts at my personal blog here and here. See also: Chomsky interviewed by Ali G. Update: Carl Zimmer writes about the paper, with quotes from many, in the May 2 NYT. Also, Chris at MixingMemory has a post on the topic here.
Schoolchildren share the Nordic look, say genetics researchers. The story looked like a hoax to me, but you can see the original data set.
Readers of John Hawks will already have seen his account of a recent paper on the evolution of cooperation.
The details are: Mikhail Burtsev and Peter Turchin: 'Evolution of cooperative strategies from first principles', Nature, vol. 440, 20 April 2006, 1041-1044. Here is the abstract: One of the greatest challenges in the modern biological and social sciences is to understand the evolution of cooperative behaviour. General outlines of the answer to this puzzle are currently emerging as a result of developments in the theories of kin selection, reciprocity, multilevel selection and cultural group selection. The main conceptual tool used in probing the logical coherence of proposed explanations has been game theory, including both analytical models and agent-based simulations. The game-theoretic approach yields clear-cut results but assumes, as a rule, a simple structure of payoffs and a small set of possible strategies. Here we propose a more stringent test of the theory by developing a computer model with a considerably extended spectrum of possible strategies. In our model, agents are endowed with a limited set of receptors, a set of elementary actions and a neural net in between. Behavioural strategies are not predetermined; instead, the process of evolution constructs and reconstructs them from elementary actions. Two new strategies of cooperative attack and defence emerge in simulations, as well as the well-known dove, hawk and bourgeois strategies. Our results indicate that cooperative strategies can evolve even under such minimalist assumptions, provided that agents are capable of perceiving heritable external markers of other agents. I have posted previously on the subject of cooperation, or the related problem of altruism, and I am sceptical about the realism of much of the game-theoretical work: for some criticisms see here. However, the paper by Burtsev and Turchin does have some promising features. One is that it allows for a wider range of strategies than most previous models. Strategies are allowed to evolve by natural selection from more basic behavioural variables. The other important feature is that individuals are allowed to recognise the degree of 'similarity' of other individuals to themselves. It is found that this influences their behaviour. In this model the only source of 'similarity' is genetic relatedness, but the authors say that they intend to extend their approach to take account of cultural transmission. As I've probably said before, I think there is a crying need for someone to give an expert but non-technical presentation of all this recent work to a wider public. (I'm thinking of something like Matt or Mark Ridley's level of popularisation.) The 'someone' would need to be an expert in game theory (so it ain't me, babe), knowledgeable about both anthropology and evolutionary theory, a very clear writer, and capable of being impartial between the various models and theories, as well as being objective about the whole game-theoretical approach (which excludes many of the direct participants in this work!) John Maynard Smith could have done it, but alas, it won't be him. |
  Razib's Home Page GNXP Archives Interviews Blogroll Principles of Population Genetics Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism
Archives
July 2005 August 2005 September 2005 October 2005 November 2005 December 2005 January 2006 February 2006 March 2006 April 2006 May 2006 June 2006 July 2006 August 2006 September 2006 October 2006 November 2006 December 2006 January 2007 February 2007 March 2007 April 2007 May 2007 June 2007 July 2007 August 2007 September 2007 October 2007 November 2007 December 2007 January 2008 February 2008 March 2008 April 2008 May 2008 June 2008 July 2008 August 2008 September 2008 October 2008 November 2008 December 2008 January 2009 February 2009 March 2009 April 2009 May 2009 June 2009 July 2009 August 2009 September 2009 October 2009 November 2009 December 2009 January 2010 Hello Movable Type archives August 11,2002 August 18,2002 August 25,2002 September 01,2002 September 15,2002 October 20,2002 December 08,2002 December 22,2002 December 29,2002 January 05,2003 January 12,2003 January 19,2003 January 26,2003 February 02,2003 February 09,2003 February 16,2003 February 23,2003 March 02,2003 March 09,2003 March 16,2003 March 23,2003 March 30,2003 April 06,2003 April 13,2003 April 20,2003 April 27,2003 May 04,2003 May 11,2003 May 18,2003 May 25,2003 June 01,2003 June 08,2003 June 15,2003 June 22,2003 June 29,2003 July 06,2003 July 13,2003 July 20,2003 July 27,2003 August 03,2003 August 10,2003 August 17,2003 August 24,2003 August 31,2003 September 07,2003 September 14,2003 September 21,2003 September 28,2003 October 05,2003 October 12,2003 October 19,2003 October 26,2003 November 02,2003 November 09,2003 November 16,2003 November 23,2003 November 30,2003 December 07,2003 December 14,2003 December 21,2003 December 28,2003 January 04,2004 January 11,2004 January 18,2004 January 25,2004 February 01,2004 February 08,2004 February 15,2004 February 22,2004 February 29,2004 March 07,2004 March 14,2004 March 21,2004 March 28,2004 April 04,2004 April 11,2004 April 18,2004 April 25,2004 May 02,2004 May 09,2004 May 16,2004 May 23,2004 May 30,2004 June 06,2004 June 13,2004 June 20,2004 June 27,2004 July 04,2004 July 11,2004 July 18,2004 July 25,2004 August 01,2004 August 08,2004 August 15,2004 August 22,2004 August 29,2004 September 05,2004 September 12,2004 September 19,2004 September 26,2004 October 03,2004 October 10,2004 October 17,2004 October 24,2004 October 31,2004 November 07,2004 November 14,2004 November 21,2004 November 28,2004 December 05,2004 December 12,2004 December 19,2004 December 26,2004 January 02,2005 January 09,2005 January 16,2005 January 23,2005 January 30,2005 February 06,2005 February 13,2005 February 20,2005 February 27,2005 March 06,2005 March 13,2005 March 20,2005 March 27,2005 April 03,2005 April 10,2005 April 17,2005 April 24,2005 May 01,2005 May 08,2005 May 15,2005 May 22,2005 May 29,2005 June 05,2005 June 12,2005 June 19,2005 June 26,2005 July 03,2005 July 17,2005 August 07,2005 Blogspot archives June 2002 July 2002 August 2002 September 2002 October 2002 November 2002 December 2002
10 questions for....
Parag Khanna James Flynn Jon Entine Gregory Clark György Buzsáki Heather Mac Donald Bruce Lahn A.W.F. Edwards Luigi Luca Cavalli-Sforza Joseph LeDoux Matthew Stewart Charles Murray James F. Crow Adam K. Webb Justin L. Barrett David Haig Judith Rich Harris Ken Miller Dan Sperber Warren Treadgold Armand M. Leroi John Derbyshire
Blogs
The GiveWell Blog Your Religion Is False Colby Cosh Steve Hsu Audacious Epigone Catallaxy Files Inductivist 2 Blowhards Genetic Future Agnostic Steve Sailer Dienekes Derek Lowe Razib Khan Razib at Comment is Free Secular Right Glenn Reynolds Jim Miller Kevin McGrew John Hawks Peter Fost Randall Parker Less Wrong Charles Murray Carl Zimmer EconLog Marginal Revolution
Principles of Population Genetics
Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism   Policies Terms of use © http://www.gnxp.com Razib's total feed: |
{kind=link}