
|
Monday, December 15, 2008
 In Jason's post on the distributions of hair and eye color, it looks like women are claiming their hair is lighter than it is. The sex differences are the opposite of what is found when the hair is rated by others. Women are lying because they think it makes them look better. In Jason's post on the distributions of hair and eye color, it looks like women are claiming their hair is lighter than it is. The sex differences are the opposite of what is found when the hair is rated by others. Women are lying because they think it makes them look better.If they're going to misrepresent their hair color, they're likely to dye it for the same reason. Whether it is to participate in the fashion for blonds in their age cohort or to cater to men in their cohort (there's evidence that younger audiences aren't as captivated by light hair), older women should lighten their hair more. It's like a boob job. I compared age and hair color data from Playboy Playmates of the Month. I only distinguish between blonds and non-blonds (though there are a few tough calls who I label intermediate). I can't look at age by year since some years have few points. I've grouped the playmates into three ranges: 16 - 20 (n = 210), 21 - 24 (n = 329), and 25 - 35 (n = 105). I chose these groups since each has lots of points. I had to include everyone 25+ in one group, or statistical tests could not have been done. In any case, these groups also correspond pretty nicely to three phases of a female's reproductive career: 1) the high school or college girl who doesn't have to try at all to look presentable; 2) the early-mid 20-something whose waistline has begun expanding; and 3) the 25+ woman who should have had a child. Here is how the female body shape changes across the lifespan. So here is a plot of the proportion of each age group that is blond, along with 1 standard error on either side: 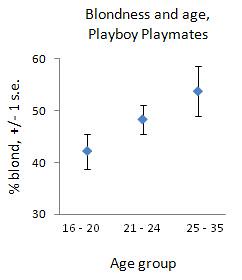 The youngest group (where blonds are 42.1%) is statistically significantly different from the oldest group (where blonds are 53.7%), and marginally significantly different from the middle group (where blonds are 48.3%). The middle and oldest groups are not significantly different. (See Appendix for the gory details.) I conclude that the 16 - 20 year-old playmates had such flawless and tight skin, clear large eyes, gravity-defying breasts and buttocks, etc., that blond hair wouldn't add much. Already by their mid-20s, women's looks have just passed their peak, so that they're probably more likely to dye their hair, although the difference may be illusory. But certainly by 25, the rest of their face and body couldn't compete with those of younger girls, so they begin dying their hair blond to distract the audience from that. (Getting fake boobs would probably show a similar age-related trend.) It's like how restaurants scam the elites by making shitty chicken sandwiches but then tossing on a bit of pesto to make it seem exotic and totally worth $15. There's another prediction of this idea: if playmates were to be drawn increasingly from older women, they should become blonder too. I've shown that the average playmate has gotten older since a low during most of the 1960s, and in the second link in this post I show that they've also gotten blonder. Indeed, the Spearman rank correlation between a year's average age and average blondness of playmates is +0.35, two-tailed p = 0.006. Taken as a whole, these differences suggest that, even if it isn't as strongly related to aging as is buying moisturizers and skin-firming lotions, lightening the hair is one way that aging women cope with their declining attractiveness. But if blond hair enhanced attractiveness to the same degree across all ages, then age would not predict the percent of women who dye their hair -- younger girls would do so just as eagerly, as with washing and styling their hair in the morning (a huge boost over a rat's nest). It seems, then, that blondness yields diminishing returns in attractiveness -- I mean, you can't really picture teen star Selena Gomez having to dye her hair. Or for that matter Audrey Hepburn, Jean Shrimpton, Penelope Cruz, or Monica Bellucci bleaching their hair at any age! Appendix I ran an F-test for equality of variances between two samples, one test for each pairwise comparison. None of the three groups had significantly different variances. I then ran a two-proportion z-test (equal variances), one test for each pairwise comparison. The p-values for the tests are: 0.080 (youngest vs. middle), 0.025 (youngest vs. oldest), and 0.164 (middle vs. oldest). These are one-tailed because the prediction was that increasing age should increase the percent who dye their hair or need blond hair to make up for having less attractive skin, breasts, etc. It's like expecting the percent of women with fake boobs and facelift surgery to increase as age increases. But I made three comparisons, and the more comparisons you make, the more likely you are to find apparently low p-values just by chance. No one really knows how to deal with adjusting the critical p-value (alpha) when you're making multiple comparisons. So rather than futz around with the many theoretical corrections to alpha, I decided to take an empirical attack. I wrote a program in Python that took all the playmates and separated them into three groups, each one having the same size as the three groups I created based on age. But instead of deterministically using age to sort them, I sampled them at random without replacement to fill the first group, then the second group, with the rest going into the third. Because "group 1," "group 2," and "group 3" were formed randomly, the proportion of blonds shouldn't be too different between them. I performed the same two-proportion z-test (equal variances) as before, one for each pairwise comparison. I simulated this process 10,000 times, and then took the fraction across all runs that the z-statistic was at least as large as the observed z-statistic, doing so for each of the three group comparisons. These empirical p-values are: 0.0813 (youngest vs. middle), 0.0226 (youngest vs. oldest), and 0.1564 (middle vs. oldest). Therefore, in this case, correcting alpha for multiple comparisons would have been pointless, perhaps because I only made three rather than three thousand comparisons. Labels: anthropometry, babes and hunks, Pigmentation
Saturday, December 13, 2008
So sayeth Aggro in the thread down below:
"They should have measured eye and hair color -- we don't have any representative data! Seriously, they'll take extra long to measure all kinds of weird things that only an anthropometer would know of, but not eye and hair color." I too have previously lamented this odd failure in easy measurement. A literature search had me coming up short for an adequate published sample of American eye and hair color. The best estimate I could cobble together from several small studies was that about 25% of American whites were blond. But, Ho Ho!, the National Longitudinal Survey of Youth is online and carries these simple treasures within its bosom. The following hair and eye color information was self-reported in 1985 by a representative sample of those born between 1957-1965 (ages 20-28; currently 43-51). I've included blacks and Hispanics for the gender breakdown: 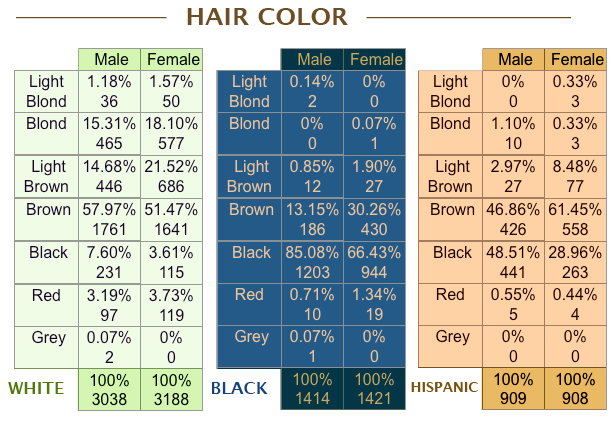
Labels: anthropometry, human biodiversity, NLSY, Pigmentation
Friday, September 19, 2008
Thank god for animals and their resemblance to humans to elucidate general patterns and relationships. Missense Mutation in Exon 2 of SLC36A1 Responsible for Champagne Dilution in Horses:
The purpose of this study was to uncover the molecular basis for the champagne hair color dilution phenotype in horses. Here, we report a DNA base substitution in the second exon of the horse gene SLC36A1 that changes an amino acid in the transmembrane domain of the protein from threonine to arginine. The phenotypic effect of this base change is a diminution of hair and skin color intensity for both red and black pigment in horses, and the resulting dilution has become known as champagne. This is the first genetic variant reported for SLC36A1 and the first evidence for its effect on eye, skin, and hair pigmentation. So far, no other phenotypic effects have been attributed to this gene. This discovery of the base substitution provides a molecular test for horse breeders to test their animals for the Champagne gene (CH).  Is horse color a big deal in terms of value? I wonder what the reason why there are so many horse pigmentation papers as opposed to a cheaper multi-colored animal like dog or cat. Is horse color a big deal in terms of value? I wonder what the reason why there are so many horse pigmentation papers as opposed to a cheaper multi-colored animal like dog or cat.Related: White horses and blonde humans: a genetic connection? KITLG makes you whiter. Labels: Genetics, Pigmentation
Thursday, August 21, 2008
Reading up on the MC1R, I came across this nice summary of work I mentioned before on the genetics of coat color in the dog. The summary includes the figure below, which is a pretty intuitive illustration of a few of the genes involved in pigment type-switching (ie. production of eumelanin versus pheomelanin). The caption:
Production of yellow versus black pigment in dogs is controlled by three genes: Mc1r, Agouti, and CBD103. Dogs carrying wild-type alleles for all three genes have a yellow coat resulting from Agouti antagonism of Mc1r signaling in melanocytes (yellow Great Dane, top). Dogs carrying a loss-of-function mutation at Mc1r have a yellow coat, regardless of their genotype at Agouti or CBD103 (yellow Labrador Retriever, middle). Dogs carrying wild-type alleles for Mc1r and Agouti, together with the dominant black allele of CBD103 (KB) have a black coat resulting from the interaction between a beta-defensin and Mc1r (black Curly Coated Retriever, bottom). 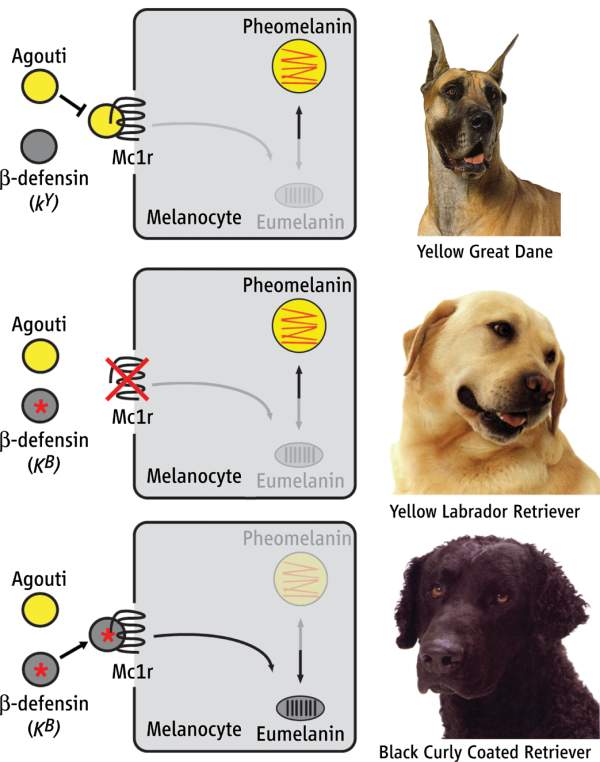 Labels: Genetics, Pigmentation
Sunday, August 17, 2008
In many vertebrates, there is an association between pigmentation and behavior. One potential reason for this is that genes influencing pigmentation also have pleiotropic effects on other traits, including behavior. A recent paper in Trends in Ecology and Evolution lays out this hypothesis:
In vertebrates, melanin-based coloration is often associated with variation in physiological and behavioural traits. We propose that this association stems from pleiotropic effects of the genes regulating the synthesis of brown to black eumelanin. The most important regulators are the melanocortin 1 receptor and its ligands, the melanocortin agonists and the agouti-signalling protein antagonist. On the basis of the physiological and behavioural functions of the melanocortins, we predict five categories of traits correlated with melanin-based coloration. A review of the literature indeed reveals that, as predicted, darker wild vertebrates are more aggressive, sexually active and resistant to stress than lighter individuals. Pleiotropic effects of the melanocortins might thus account for the widespread covariance between melanin-based coloration and other phenotypic traits in vertebrates. This is clearly far from gospel truth; the authors are laying out the plausibility of this hypothesis and a framework for further exploration. The hypothesis is that higher levels of the molecules that bind the melanocortin receptors (the melanocortins and agouti proteins) lead to both darker pigmentation as well as pleiotropic effects in other tissues (I've mentioned before some of the effects of messing with these receptors in sexual behavior and metabolism). Analysis of the way pigmentation and various other traits vary in mouse models leads to results consistent with this hypothesis. A corollary of this argument is that in vertebrates where pigmentation is controlled downstream of the melanocortins (ie. at, or further downstream of, MC1R), this correlation between pigmentation and other traits should not be consistently true. For this reason, the authors argue that humans should be exempt. However, they may be unaware that some difference in pigmentation both between and within populations is controlled by ASIP, a protein that binds MC1R, acting as an antagonsist for melanocortin binding. Humans, then, could be an ideal test case for the hypothesis--do phenotypes like aggression map to ASIP like pigmentation does? However, the unpalatable nature of this question makes it rather unlikely to be pursued in humans. Labels: Genetics, Pigmentation
Sunday, July 06, 2008
Maxim's audience prefers brunettes; distribution is bimodal
posted by
agnostic @ 7/06/2008 03:10:00 PM
While showing that the super-popularity of blonds is recent, I saw an apparent reversal of the upward trend around 2000, suggesting that perhaps Playboy readers are becoming fatigued by blonds. To get a better feel for what the younger generations prefer, let's look at Maxim magazine (US edition), whose average reader is 27.5 years old (by contrast, the average Playboy reader is 32.5). Maxim is the contemporary counterpart to Playboy -- it's widespread on college campuses, and is what horny dudes are likely to leaf through to ogle hot babes. They also have roughly the same circulation -- about 2.5 million. For those in a rush, I've boldfaced all key results.
Audacious Epigone and I have both done analyses on the Maxim Hot 100 lists for recent years (see here and here). But these smell of "lists just for the fun of making lists" or "lists to get people arguing," and the fact that A.E.'s results and mine varied so much despite analyzing consecutive years supports that idea. However, there are at least two datasets that are surely more shaped by audience demand than the editors' whims: the girls who appear on the cover to lure the reader into purchasing it (they are featured prominently inside as well), and the girls who readers vote as being the hottest out of a pool of "Hometown Hotties" nominees. First, I looked at all covers of Maxim from 1997 to present, excluding only two issues that did not show people, which yielded 126 covers. To be more fine-grained than before, I coded hair color as 1 = light blond, 2 = dark blond, 3 = light brown, 4 = dark brown, and set aside 0 for redheads. If there were multiple girls on a cover, or if an issue had multiple covers available, I took the average of all girls for that issue. For a few, it was too close to call, so I coded the girl halfway between two categories. If the cover was ambiguous, I looked at the full photo shoot through a Google Images search. There was no significant trend in blondness over the past 11 years. To determine the average hair color, I re-coded redheads as 2.5 (there were only 2 of 126, so this choice doesn't really make a difference). The average Maxim cover girl scores 2.8 -- light brown. To determine the frequency distribution, I binned girls into light blond, dark blond, light brown, and dark brown, with redheads going into the light brown bin. Some data-points were not integers, so I used both conventions for rounding the numbers with .5 ("up" and "down"). It turned out to make almost no difference. Here is the distribution using "rounding up": 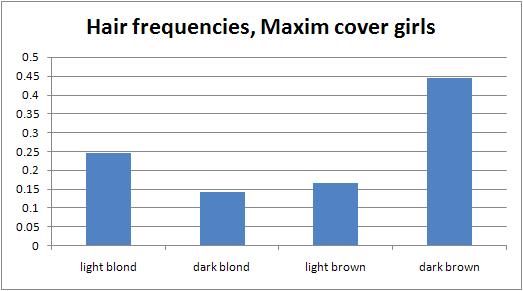 Light blonds and dark browns are overrepresented, while the intermediate colors are underrepresented. To test this, I used published data on hair color frequencies and took the Dutch values instead of the Icelandic ones, since Americans must resemble the former more than the latter. Because the published estimates have only one intermediate category, I had to merge the dark blond and light brown categories together. I put redheads in the intermediate category since otherwise I wouldn't be able to do a chi-squared test (an expected number would be too small). For rounding up, chi-squared = 24.1 (p less than 0.0001, df = 2); and for rounding down, chi-squared = 27.4 (p less than 0.0001, df = 2). So, the discrepancy between Maxim cover girls and the general population is no fluke. But are light blonds and dark browns equally overrepresented? No: depending on the rounding convention, light blonds are 25-29% more common than we'd expect, whereas dark browns are 63-66% more common than we'd expect. Together with the average cover girl being light brown, I conclude that Maxim readers respond more to women with dark hair, although there is a sizable minority that prefers light hair. Second, I did a similar analysis on the finalists in Maxim's Hometown Hotties contest from 2003 to 2007. For 10 weeks each year, Maxim staffers scour the country to photograph 100 local hotties per week. Of these 100, Maxim readers vote online to determine 10 semi-finalists and 1 finalist for that week. There is no way I'm looking through 5000 pictures to see what all the contestants look like, and the 5 winners are too small of a sample. The 50 finalists seem like enough data to get a good picture. (Someone else can analyze all 500 semi-finalists.) Indeed, the results are virtually identical to the cover girl results, which shows that both datasets are reliably measuring the same thing. The methods are as before. Here is the distribution of hair types among Hometown Hotties finalists: 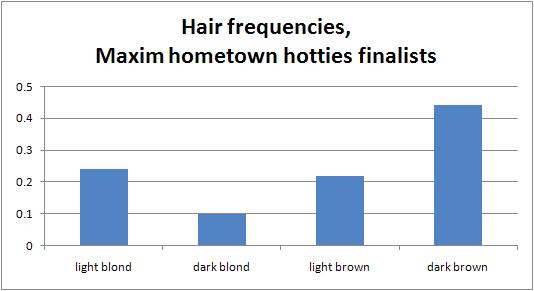 Once more, light blonds and dark browns are overrepresented, while intermediate colors are underrepresented. For rounding up, chi-squared = 10.3 (p = 0.0058, df = 2); for rounding down, chi-squared = 13.1 (p = 0.0014, df = 2). These results are no fluke. As before, though, dark browns are more overrepresented than light blonds: by 57-64% compared to 22-52%, respectively, depending on the rounding convention. (The convention for rounding didn't make much of a difference overall in these data either, but since the sample size is less than half that of the cover girl data, it introduces more uncertainty.) Replicating the cover girl results, the average Hometown Hotties finalist scores 2.8 -- light brown. I conclude what I did in the cover girl case. To see how closely the two datasets agree with each other, I did a chi-squared test for the observed values in one, using the other's frequencies as the expected ones. Taking the cover girl frequencies as expected, the hometown hotties data are no different (chi-squared = 0.025, p = 0.9875, df = 2). The same holds for the other way around (chi-squared = 0.068, p = 0.9666, df = 2). That is for rounding up, but rounding down produced p-values above 0.5 as well. I conclude that both datasets measure the same thing -- audience preferences. As a final anecdote in support of the bigger picture, consider the Miss Maxim girls. Although about 1/2 of the 24 countries could have easily supplied a blond, only 1/6 actually did. The girls from Belgium, England, and so on, look quite different from the average Belgian, Englishwoman, etc. Clearly, among Maxim's horndog audience, dark hair rules. What is causing these two key results -- that Maxim readers prefer brunettes, and that the distribution is bimodal? I think brunettes are just more exciting on the level of physiological arousal, so the younger -- and therefore the randier -- the audience is, the more they will prefer dark hair. When Playboy's circulation was growing exponentially in the 1960s, it featured hardly any blonds and mostly brunettes. As its average reader has become older, its Playmates have become blonder. Lighter hair is correlated with behavioral inhibition (see here), so it could also be that dark-haired girls get the blood pumping more because they appear more flirtatious. Or it may be a pure fashion trend -- digging blonds is what your father's generation did, so you set yourselves apart by tacking up pictures of Mila Kunis and Vanessa Minnillo on your wall. As for the bimodal nature of the distribution, this probably reflects supply meeting demand: the audience's preferences are likely bimodal, with a majority preferring brunettes and a minority preferring blonds. Guys respond better to the exaggerated version of their tastes, and that drives up the fraction of light blonds and dark browns, in the same way that among porn stars you see an inflated fraction of women with large breasts or large rumps. Labels: babes and hunks, Pigmentation
Sunday, June 29, 2008
 Long-time readers know that one of my beliefs that I'll stop at nothing to prove is that blond women are not sexier than brunettes, whatever other appeal they obviously have for many men. Point-estimates of the current popularity of blond hair neglect the fact that standards of beauty can change over time -- within boundaries, to be sure, but still. Perhaps we only live in a blond-obsessed world today, while brunettes may have ruled in the past. Indeed, I will show just that. Furthermore, the shift toward blonds parallels several other shifts toward a more masculine ideal of female beauty since the early / mid-1960s. Long-time readers know that one of my beliefs that I'll stop at nothing to prove is that blond women are not sexier than brunettes, whatever other appeal they obviously have for many men. Point-estimates of the current popularity of blond hair neglect the fact that standards of beauty can change over time -- within boundaries, to be sure, but still. Perhaps we only live in a blond-obsessed world today, while brunettes may have ruled in the past. Indeed, I will show just that. Furthermore, the shift toward blonds parallels several other shifts toward a more masculine ideal of female beauty since the early / mid-1960s.The data come from Playboy Playmates of the Month ("Playmates") from 1954 to 2007. We need to look at sources that pander to popular demand in sexual tastes, which excludes runway fashion magazines (not used by males for fantasy purposes) as well as data on high-ranking Hollywood actresses (who are esteemed only in part based on their looks). We also need comparable data that stretch over decades, and that provide us with many data-points for each year -- in a worst case scenario, we might look at something like Miss Universe winners, but estimating the value of blond-obsession for a given year with only a single data-point is hardly ideal. Playmates, though, yield 12 data-points per year. In the name of scientific discovery, I looked at pictures of every Playmate [1], and coded her hair color as either 1 for blond or 0 for non-blond. Dark blonds counted as blond, light browns as non-blond. Redheads counted as blond if they had very fair, strawberry blond hair, and as non-blond otherwise. The point is not to measure the popularity of the full spectrum of hair colors -- just blondness. A small handful of Playmates had several hair colors within the single issue they appeared in. I coded these as 0.5 because their pictures were split pretty evenly between blond and non-blond hair -- maybe due to wigs, I don't know. I then took the fraction of blonds in a given year and plotted these over time. Here is the raw scatter-plot, together with 3-year and 7-year moving averages that smooth it out: 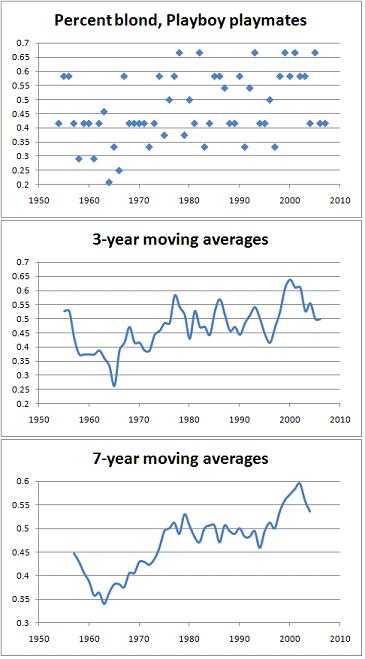 The scatter-plot suggests an increasing trend, and this is true: Kendall's tau for the correlation between year and percent blond is +0.27 (p = 0.01, two-tailed). [2] However, because each year's value can only take on roughly 12 values (1 / 12, 2 / 12, etc.), there are a lot of tied years, which may underestimate the true correlation. Kendall's tau for the correlation between year and the 3-year moving average of percent blond is +0.47 (p = 2 x 10^(-6), two-tailed), and is +0.64 (p = 2 x 10^(-10), two-tailed) when the 7-year moving averages are used. Using a moving average gives us a better idea since they can take on far many more values, and so produce fewer ties. Whichever one we choose, it is clear that blonds have increased quite a bit in popularity over the decades. At the same time, the trend is clearly not linear: there is a decrease in blond-obsession at least from the mid-1950s, when the data begin, to the early / mid-1960s. There follows an increase, and an apparent reversal since the turn of the millennium -- please god, let it be so. This looks periodic, like a fashion cycle. In trying to account for this trend, we should try to be as general as possible. What other trends in female beauty show an increase after the early 1960s? I didn't look at other aspects of the Playmates, but someone else has tabulated data on Playmates of the Year from 1960 to 2006 -- again, estimating the popularity of some trait in a given year based on a single data-point is worst-case, but I'm relying on it here because I've already spent enough time collecting hair color data. The links in footnote 1 provide all the anthropometric data, though, so if you want to collect an analyze it, we will link to your analysis. I calculated the Waist-to-Hip Ratio and BMI of Playmates of the Year from the above data, and Kendall's tau for the correlation between year and WHR is +0.53 (p = 4 x 10^(-7), two-tailed), while between year and BMI it is -0.24 (p = 0.02, two-tailed). So, these sex symbols are increasingly losing their feminine hourglass shape and fatty softness -- nearly all BMI points are below 20, so it's not like they used to be fat but are now healthy. They are also getting taller: Kendall's tau is +0.31 (p = 0.004); and smaller in the chest: Kendall's tau is -0.35 (p = 0.001). Someone else has also done an analysis of Miss America winners, and the exact same trends emerge there as well (see his graphs). The common factor of all these trends is that the ideal of female beauty has become increasingly masculinized. Recall that males are more likely to be blond, so the hair color trend is part of the larger masculinizing trend. I didn't look at eye color, but if it's part of the overall trend, the earlier Playmates should be less blue-eyed than later ones, as blue eyes are also more typical of males. Skin color would be tougher to analyze; if it's part of the same trend, it should get darker over time. Anecdotally, these two guesses seem to be true, but someone should look at the data to check. It therefore appears that a preference for blonds should also correlate with a preference for taller and less curvy women. Again, someone else can look that up in the psychology literature and post in the comments. But the words "tall," "thin," and "blond" usually co-occur, don't they? Whatever appeal such women have, raw sex appeal is unlikely to be among the top reasons. Blond hair correlates with something like introversion, and that makes sense since men on average are more introverted than women. So, maybe guys start digging blonds when they become more marriage-minded, or if they are inveterate monogamists. A blond will be less likely to be bouncing off the walls and being constantly out and about in search of social stimulation. Bang a brunette, bank on a blond? It would fit with the trend toward lower sluttiness in recent times, which we expect to weed out the sex kitten types from popular culture. This suggests that dark hair is part of that highly sexualized image -- something that was always obvious to everyone but the blond-lovers. [1] For years 1954 to 1992, I used this source that contains the full shoot for each Playmate, and for 1993 to 2007, Playboy's official website (if the single picture available on the Playboy site was ambiguous, I did a Google image search to get a better idea). [2] You can easily calculate Kendall's tau with this website, which I used here. Labels: babes and hunks, Pigmentation
Monday, May 19, 2008
Pigementation is turning out to be one of the most tractable phenotypes for genome-wide association studies-- a new paper from Decode identifies another couple loci that influence various aspects of hair and skin pigment. These add to the growing list of pigmentation genes in Europeans--I've not done the math, but these loci must account for some sizeable chunk of the total genetic variance in these traits.
Why has pigmentation been so amenable to mapping, while other traits like height, weight, or most diseases hover at about 2-3% of the variance explained? One possibility is that it's because pigmentation has been subject to strong recent selection--theory predicts that the initial moves toward an new fitness optimum will be loci of large effects, while the later moves will be smaller. Perhaps since this selective pressure has been so recent, we're still picking up those initial mutations of large effect that are still segregating in the population? Labels: Genetics, Pigmentation
Saturday, May 17, 2008
A Genome-Wide Association Study Identifies Novel Alleles Associated with Hair Color and Skin Pigmentation:
It has been a longstanding hypothesis that human pigmentation is tightly regulated by genetic variation. However, very few genes have been identified that contain common genetic variants associated with human pigmentation. We scanned the genome for genetic variants associated with natural hair color and other pigmentary characteristics in a multi-stage study of more than 10,000 men and women of European ancestry from the United States and Australia. We identified IRF4 and SLC24A4 as loci highly associated with hair color, along with three other regions encompassing known pigmentation genes. Further work is needed to identify the causal variants at these loci. Improved understanding of the genetic determinants of human pigmentation may help identify the molecular mechanisms of pigmentation-associated conditions such as the tanning response and skin cancers. There are four regions because areas around HERC2/OCA2 and MAPT showed signals. MAPT is also known as AIM1 and SLC45A2, so this makes 3 genes of the potassium-dependent sodium/calcium exchangers implicated in pigmentation (the other is SLC24A5 obviously). They adjusted for the components of genetic variation so as not to be confounded by population stratification (i.e., there was some ethnic variation among their whites and so you don't have a random mating population). It's in PLOS; you can read the whole thing, etc. Related: Why white people are so colorful!. Sandy also comments. Labels: Genetics, Pigmentation
Tuesday, May 06, 2008
In the comments here, rosko points me to a study on the effects on MC4R, a gene implicated in natural variation in human weight, on pathways involved in sexual function. It's well known, of course, that genetic pathways can be involved in multiple physiological processes--in particular, signaling pathway can generate many different phenotypes depending on what the downstream target of the signal is.
The effects of MC4R simulation in humans are, as rosko comments, kind of interesting: Methods. Ten subjects were enrolled in a double-blind, placebo-controlled, crossover study. Melanotan II (0.025 mg/kg) and vehicle were each administered twice by subcutaneous injection; real-time RigiScan monitoring and a visual analog were used to quantify the erections during a 6-hour period. The level of sexual desire and side effects were recorded with a questionnaire.I wondered what a "Rigiscan" is--find out here. Hypothetically, one could test whether natural variation in sexual behavior in humans is also affected by MC4R polymorphism, though I can't imagine that being a particularly fun study to carry out (one for agnostic's new series? 23andme + free time = association studies about erections). This reminds of the MC1R story about increased pain sensitivity in redheads in the vague sense that both involve melanocortin receptors and pleiotropy. Labels: Genetics, Pigmentation
Monday, May 05, 2008
For many years, Grey Squirrels (an introduced North American species) has been driving out the indigenous Red Squirrel over most of mainland Britain. But now it is reported that a mutant black variety of the Grey Squirrel is threatening to displace the Greys. Apparently, the black ones have higher testosterone levels, are more aggressive, and more attractive to the lady squirrels. (Don't worry, our White Nationalist readers, this isn't a parable. I think.)
Joking apart, the real interest of this is that it seems to be a case of a single mutation with a relatively conspicuous phenotypic effect having a strong evolutionary advantage, somewhat contrary to Darwin/Fisher orthodoxy. There is of course another example in the case of industrial melanism. Labels: Pigmentation
Wednesday, February 27, 2008
How Skin Color Is Determined:
In 2005 researchers identified a gene called SLC24A5 as a key determinant of skin color. Rebecca Ginger and colleagues now confirm that the protein product of this gene (NCKX5) is an ion exchanger; it exchanges sodium for calcium across a membrane, regulated by potassium. But unlike other NCKX proteins, they found that NCKX5 is not present on the cell surface, but internally in a compartment known as the trans-Golgi network. This compartment is where new proteins and vesicles are processed, modified and sorted. Related: SLC24A5 and skin color. Labels: Genetics, Pigmentation
Saturday, February 02, 2008
I've been blogging the HERC2/OCA2 story a fair amount. It seems this genomic region is the locus of main effect for variation of eye color in Europeans, in particular blue vs. non-blue eyes. But I also pointed out that this locus has also been connected to variation in skin color, and while that variation is additive in effect, the variation on eye color exhibits strong dominance/recessive dynamics. My inference here is that it is more plausible that selection occurred on skin color, while eye color was a tissue specific expression pattern which emerged as a byproduct. Peter Frost has an objection to this:
The correlation between eye color and skin color may simply be an artefact of geographic origin. Europeans vary clinally for both eye color and skin color along a north-south and west-east gradient, so if the pool of subjects is geographically heterogeneous you will almost certainly get a correlation between eye and skin color. But this doesn't prove a cause and effect relationship. Fair enough. Spurious associations driven by cryptic population substructure is one of the main reasons Structure was developed. I responded to Peter here, here and here. The short of is that I don't know of any analysis within an admixed population like African Africans, which would settle the matter, but there are plenty of other points which would suggest that we should look at the skin color trait (and, to be fair, if substructure exists at the level of British Isles origin samples we really need Strucure!). But there was something that has been bothering me: eye color difference exhibits a lot of dominance/recessive dynamics in expression. The skin color data here does not, and aside from KITLG (which is dominant for light skin) all the other loci seem additive and independent (the report of epistatic effects here & there don't seem reproduced very often). One of the main reasons that I am favoring a skin color model as the phenotype driving selection is that if it is additive it is exposed to selection immediately at low frequencies. In contrast, recessive traits at low frequencies have the problem that most copies of the allele which increases fitness are still in heterozygotes which mask them from selection. It came to my mind that the different assumptions about dominance would matter in terms of long term evolutionary dynamics and how that would be realized in terms of results from tests for selection. So I found this paper, Directional Positive Selection on an Allele of Arbitrary Dominance. It says: ...fixation of a beneficial allele leaves a signature in patterns of genetic variation at linked neutral sites. If this signature is well characterized, it can be used to identify recent adaptations from polymorphism data. To date, most models developed to characterize the effects of positive directional selection (termed "selective sweep") have assumed that the favored allele is codominant. In other words, if the fitnesses of the three genotypes are given by 1, 1 + sh, and 1 + s (where s is the selection coefficient), then h = 1/2.... For skin color h would be 1/2 for HERC2/OCA2, it has half the effect on the trait value. Assuming proportional selection based on the character value two copies would be better than one copy which would be better than no copies. In contrast, for eye color the h would be between 0 and 1/2, and probably closer to 0 because of predominant recessivity in expression for blue eyes. That means the fitness of those with one blue eye copy would be much closer to those with no blue eye copies than those with two; to the homozygote recessives would go all the benefit. On to the results: ...when h is small, most of the sojourn time is when the allele is at low frequency in the population. During this phase, the allele will have the opportunity to recombine onto other backgrounds. In other words, the favored allele will tend to increase in frequency on multiple backgrounds, preserving more of the diversity that existed when it first arose. In contrast, for dominant alleles, most of the sojourn time is spent at higher frequency, when there is less opportunity for the favored allele to recombine onto other backgrounds. This results in a wider signature of a fixation event for larger h-values. Why the bolded parts? From A Map of Recent Positive Selection in the Human Genome: Some of the strongest signals of recent selection appear in various types of genes related to morphology. For example, four genes involved in skin pigmentation show clear evidence of selection in Europeans (OCA2, MYO5A, DTNBP1, TYRP1). All four genes are associated with Mendelian disorders that cause lighter pigmentation or albinism, and all are in different genomic locations, indicating the action of separate selective events. One of these genes, OCA2, is associated with the third longest haplotype on a high frequency SNP anywhere in the genome for Europeans.... I don't know if my connection of inferences here is valid, and the paper I originally referenced makes clear that it is important to frame these sorts of assumptions within their statistical context; just because something is less likely does not mean it is impossible. I've sent out emails about OCA2 and skin color, and will report back, but at this point I suspect that the final proof in the pudding will have to be admixture analysis in a group like African Americans. But I think the above makes it more likely that whatever was going on 10,000 years ago did not express as a recessive phenotype. Labels: Genetics, Pigmentation
Friday, February 01, 2008
The story about HERC2 & OCA2 is getting a lot of press; that is, the genetics behind how people have blue eyes. But see this in ScienceNow:
There are still large questions, though. Why did blue eyes persist? Scientists say it is difficult to see how eye color would have an environmental advantage, as skin color does. Some theories suggest that women may have played a role in driving the selection. Perhaps, Kayser says, "the females thought it more exciting to have a male with blue eyes." I already posted this before: the SNPs which are used to predict blue eyes also track skin color variation. In other words, pleiotropy. This shouldn't be a surprise, OCA2 is a pigmentation locus which in many cases doesn't exhibit tissue specific expression patterns; its name derives from the fact that some forms of albinism are associated with mutants on it. In any case, some concrete data about skin color and the OCA2 SNPs can be found in a previous paper from a research group behind one of the current publications, A Three-Single-Nucleotide Polymorphism Haplotype in Intron 1 of OCA2 Explains Most Human Eye-Color Variation. Look at table 1, and you find these data:
Let's do something with the numbers. Give fair skin a trait value of 1, medium skin 2, and olive skin 3. Then generate an average value for each genotype by weighting appropriately, and divide by the number associated with heterozygotes. This is what I get:
Looks additive for skin color, doesn't it? Since blue eyes as a trait seems to exhibit strong recessivity HERC2/OCA2 derived variants are unlikely to have initially been selected for that phenotype. It could be something besides skin color, but that is the most plausible abduction at this point from where I stand (we know that selection was powerful on the locus). Related: Genome-wide associations, HERC2 and eye color, 1 SNP to rule them & in the darkness bind them?, Why do you have blue eyes?, HERC2 & blue eye color & Danes and OCA2, blue eyes and skin color. Labels: Genetics, Pigmentation
Thursday, January 31, 2008
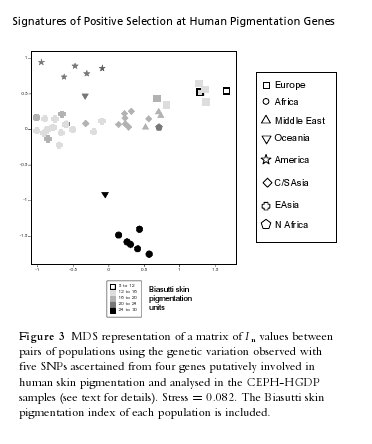 The figure to the left is from Signatures of Positive Selection in Genes Associated with Human Skin Pigmentation as Revealed from Analyses of Single Nucleotide Polymorphisms. I thought of this chart when considering the idea that the phenotypic races that we see around us might be relatively new; perhaps an artifact of recent human evolution. Look at "Oceania," those are Bougainville Islanders, from off the coast of Papua New Guinea. In the CEPH-HGDP populations the "South Asians" are from the much lighter skinned northwest fringe of the subcontinent; otherwise, I suspect you would be seeing the South Asian group moving toward the location of the Bougainville Islanders. This is not a surprising finding, earlier studies implied that very dark-skinned populations tended to exhibit a "consensus sequence" due to functional constraint; there's a reason humans are dark-skinned around the equator, and there's only one way to do it. But here's an important point: Bougainville Islanders are closer to East Eurasians than they are to other world populations in terms of ancestry. In other words, the dark-skin and the genes which confer that trait that results in an affinity between Melanesians and Africans in appearance is not a function of relatively recent common descent, but of local adaptation. Similarly, extreme dark-skinned South Asian groups are generally closer to Europeans in terms of ancestry than light-skinned East Asians. The figure to the left is from Signatures of Positive Selection in Genes Associated with Human Skin Pigmentation as Revealed from Analyses of Single Nucleotide Polymorphisms. I thought of this chart when considering the idea that the phenotypic races that we see around us might be relatively new; perhaps an artifact of recent human evolution. Look at "Oceania," those are Bougainville Islanders, from off the coast of Papua New Guinea. In the CEPH-HGDP populations the "South Asians" are from the much lighter skinned northwest fringe of the subcontinent; otherwise, I suspect you would be seeing the South Asian group moving toward the location of the Bougainville Islanders. This is not a surprising finding, earlier studies implied that very dark-skinned populations tended to exhibit a "consensus sequence" due to functional constraint; there's a reason humans are dark-skinned around the equator, and there's only one way to do it. But here's an important point: Bougainville Islanders are closer to East Eurasians than they are to other world populations in terms of ancestry. In other words, the dark-skin and the genes which confer that trait that results in an affinity between Melanesians and Africans in appearance is not a function of relatively recent common descent, but of local adaptation. Similarly, extreme dark-skinned South Asian groups are generally closer to Europeans in terms of ancestry than light-skinned East Asians.This is all pretty common sense when you think about it. But with that said skin color is a very salient trait. The skin is our biggest organ, it's a large part of what others see. Therefore, there is a natural human tendency to classify in colors. If you read the reports from Chinese delegations who were sent to investigate Cambodia they describe the natives as "black." Similarly, according to Mary Lefkowitz the ancient Greeks observed that there were the blacks of Ethiopia and those of Southern India. They also noted that both the Egyptians and North Indians were brown-skinned people ("wheat colored"). But, perhaps importantly, they often distinguished the various peoples by other characteristics (e.g., Ethiopians and Indian hair form). So on the one hand you have an nod to the importance of skin color as a criterion of perception & categorization, and on the other hand an acknowledgment that populations differ in more than color. But in the United States there are peculiar social conditions which result in problematic conflations. As everyone knows, to be very dark-skinned in the United States was identical to being of one race for a greater part of our history. Certainly there was a small Native American population, but they could be discarded from the shaping of social norms because of their low numbers. To have dark-skin was to be of African ancestry. Though there were certainly other distinguishing characteristics between those of African and European ancestry, skin color was the most visible and noticeable. It was used as the main discriminatory trait because that was all that necessary. This still persists in our folk culture when people talk about individuals "being discriminated against because of the color of their skin." Skin color connotes a racial identity. And yet you have groups like South Asians, who overlap with African Americans in complexion, but are not really"black" as we understand it. Steve Sailer has been noting for years the implicit value system highlighted by the reality that the very dark-skinned Vijay Singh is not identified as a black golfer, while the lighter-skinned (and only 1/4 African in ancestry) Tiger Woods is. Of course it doesn't work this way all the time, and South Asians are often identified as black, at least upon first impression. But the more confusing situations can also occur because of the nature of American categorizations. So tight is the correlation of non-white and "black" in the minds of some people that really peculiar characterizations can ensue. For example, in high school I had an acquaintance who would refer to myself & a Cambodian girl as black. That was understandable, we both had brown-skin. But, one day he referred to a Chinese friend of mine as black. This friend was not a dark-skinned, she had a brunette white complexion (not olive). When I queried my acquaintance about the fact that this "black" individual was probably lighter skinned than at least 1/3 of our other classmates (all of whom were white), he simply insisted that she was a "Chinese black." That was about as far as I got, obviously he couldn't express the inchoate associations within his mind between racial identity and skin color. In his world, there were whites and blacks. If someone wasn't white, that entailed that they were black. As is rather clear from the content on this weblog we are getting a good fix on the genetics of pigmentation. Not only do we know the patterns of inheritance via classical pedigree analysis, but we now have a good grasp on which regions of the genome control world-wide variation in melanin content of the skin, eye and hair. We are even beginning to understand when selection began to occur on the loci which control this variation. We have some working hypotheses of why skin color is under functional constraint, and what sort of changes might drive adaptive evolution. But all this is sometimes harder to discuss because the typical American has so many social and psychological associations between skin color and group identity. It isn't just another trait, like bristles on the back of a Drosophila, no, it is the token of one of the most significant sociological phenomena which characterize American society today. Steve will have quite a bit to blog about into the foreseeable future. Note: I suspect that the transposition of genomic knowledge to folk wisdom is easier in societies such as Brazil or India where extant phenotypic variation on this trait exhibits a larger range, much of it within families. Race and color are still very important issues, but the joints around which the perceptions are carved are more flexible and numerous. Labels: Genetics, human biodiversity, Pigmentation
Wednesday, January 30, 2008
I was doing some snooping around due some questions about the HERC2 & eye color papers I mentioned yesterday. Guess what? Earlier this month a Danish group published a similar paper, Blue eye color in humans may be caused by a perfectly associated founder mutation in a regulatory element located within the HERC2 gene inhibiting OCA2 expression. It's Open Access, so you can read it yourself. The language is a bit more stilted and hurried than the two papers I mentioned yesterday, but the basically independently confirmed the Australian group's specific finding:
In conclusion, we have identified a conserved regulatory element within intron 86 of the HERC2 gene that is perfectly associated with the brown/blue eye color in studied individuals from Denmark, Turkey and Jordan. This element had an inhibitory effect on the OCA2 promoter activity in cell cultures, and the blue and the brown alleles were shown to bind non-identical subsets of nuclear extracts. In total, all these data strongly support a model where the blue eye color in humans is caused by homozygosity of the rs12913832*G allele. Instead of just doing comparative analysis they actually tested the hypothesis in cell culture after preforming linkage & association, and seem to have come out with what you'd expect, the SNP on intron 86 of HERC2 regulates transcription at OCA2. Their Ns were a little small compared to the other two groups, but their inclusion of Middle Eastern individuals was interesting. They imply that it's a common haplotype derived from a single mutational event, presumably recently driven up in frequency by selection. Their conjecture of location and rationale aren't convincing, I'm sure commenters here could offer many more ingenious models based on historical & geographical particulars (I know the reasons proffered overlap with some of mine, but I'm a dude on a blog). I get the impression they haven't heard of Haplotter (look at the references). All that being said, at the rate that papers are being pumped out the golden age of pigmentation genetics may not have a very long shelf life (granted, that's a good thing). By the way, the gene they say has an association with hair color, RABGGTA, has been pegged as being under negative selection. Update: ScienceDaily has a summary up with a most retarded title. Labels: Genetics, Pigmentation
Monday, January 28, 2008
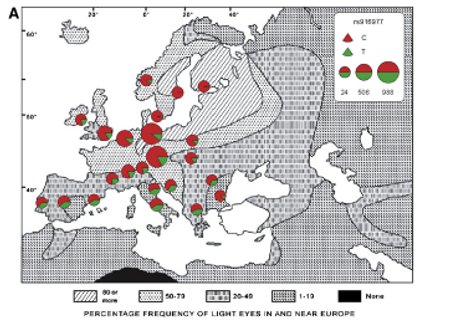 There are two new papers out in AJHG about eye color variation and genomics. Three Genome-wide Association Studies and a Linkage Analysis Identify HERC2 as a Human Iris Color Gene and A Single SNP in an Evolutionary Conserved Region within Intron 86 of the HERC2 Gene Determines Human Blue-Brown Eye Color. The second paper is an extension of the work of the Australian group which has been elucidating pigmentation relationships around OCA2 for several years now. The first paper is more interesting (to my mind) because it's the first genome-wide association study to focus on this region. I've extracted figure 6a out of the paper, you might recognize the map. I'm not surprised; go to Haplotter and enter in HERC2, it pops out as a region of selection near OCA2 (I first noticed it when checking for OCA2). As for the map, pretty cool huh? As the authors note there's a pretty good correlation between the frequency of the trait and the SNP of interest. The authors point to the north-south cline, but I am curious about the east-west one. Additionally, look at Bulgaria. I've been looking at Slavicization of the Balkans, and this is an interesting data point.... There are two new papers out in AJHG about eye color variation and genomics. Three Genome-wide Association Studies and a Linkage Analysis Identify HERC2 as a Human Iris Color Gene and A Single SNP in an Evolutionary Conserved Region within Intron 86 of the HERC2 Gene Determines Human Blue-Brown Eye Color. The second paper is an extension of the work of the Australian group which has been elucidating pigmentation relationships around OCA2 for several years now. The first paper is more interesting (to my mind) because it's the first genome-wide association study to focus on this region. I've extracted figure 6a out of the paper, you might recognize the map. I'm not surprised; go to Haplotter and enter in HERC2, it pops out as a region of selection near OCA2 (I first noticed it when checking for OCA2). As for the map, pretty cool huh? As the authors note there's a pretty good correlation between the frequency of the trait and the SNP of interest. The authors point to the north-south cline, but I am curious about the east-west one. Additionally, look at Bulgaria. I've been looking at Slavicization of the Balkans, and this is an interesting data point....Related: Dienekes has a high res map up. Note: Please be careful about taking the phenotypic clines too literally, I am to understand that there was a little extrapolation going on here and there. And of course, standard caveats on representativeness of the samples from each region and all. (Via Assman) Labels: Genetics, Pigmentation
Sunday, January 27, 2008
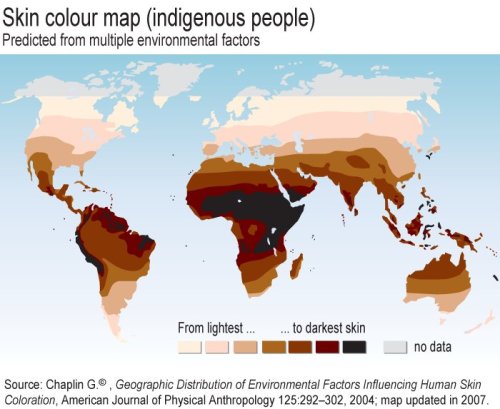 From Geographic distribution of environmental factors influencing human skin coloration: From Geographic distribution of environmental factors influencing human skin coloration:...The UVR [ultraviolet radiation] data recorded by satellite were combined with environmental variables and data on human skin reflectance in a geographic information system (GIS). These were then analyzed visually and statistically through exploratory data analysis, correlation analysis, principal components analysis, least-squares regression analysis, and nonlinear techniques. The main finding of this study was that the evolution of skin reflectance could be almost fully modeled as a linear effect of UVR in the autumn alone. This linear model needs only minor modification, by the introduction of terms for the maximum amount of UVR, and for summer precipitation and winter precipitation, to account for almost all the variation in skin reflectance..... The map above was generated from the regression analysis. Apparently it has been updated as of 2007 (received the link from a friend). It does look much better than it did in the original paper (which I have read and have a PDF copy of). Do note that the selection of peoples whose reflectance values were plugged into the model obviously matters. But I still think it's interesting the sort of predictions this map produces and how it fits with our intuitions of what the distributions should be, and the knowledge of what they are. Note the equivalent latitudes in Europe and North America, or Australia. Labels: human biodiversity, Pigmentation
Saturday, January 19, 2008
I was talking with a friend about Native American skin color. From the Canadian north down to Chile it seems that though there is variation these populations exhibit some sort of brownish shade. There are no black-skinned Amazonians, nor are there pink-skinned peoples on the Canadian Arctic. So what gives?
 First, it seems likely that Native Americans have been "native" to the New World for only around the past 10,000 years.1 A physical anthropologist once told me that the body proportions of the natives of the Amazon are still quite "Siberian," that is, they exhibit adaptations to cold weather after all these generations. And of course time is not the only parameter, Native American populations seem to have gone through a genetic bottleneck; they likely brought over very little standing genetic variation. So you have a relatively short period of time for selection to operate upon over a very limited range of trait value.2 First, it seems likely that Native Americans have been "native" to the New World for only around the past 10,000 years.1 A physical anthropologist once told me that the body proportions of the natives of the Amazon are still quite "Siberian," that is, they exhibit adaptations to cold weather after all these generations. And of course time is not the only parameter, Native American populations seem to have gone through a genetic bottleneck; they likely brought over very little standing genetic variation. So you have a relatively short period of time for selection to operate upon over a very limited range of trait value.2But this isn't persuasive to me for skin color, at least in the totality. We seem to know the genes at work now. We know that they can be selected very fast, and we know that there have been convergent dynamics across the World Island. 10,000 years is plenty of time. So perhaps the second parameter, extant genetic variation, is at work? That is, the Siberian migrants didn't bring all the genes for selection to shift them toward new adaptive optimums. For dark skin the data suggest that there is a rough consensus sequence, a constrained set of alleles across skin color genes, which produces our species' dark "Wild Type." This suite of genes probably arose when we lost our fur and became strongly pigmented to counter the negative affects of radiation, and it seems like there hasn't been any reinvention of the wheel here. Melanesian populations which are quite distant from Africans on most genes exhibit the same consensus sequences for skin color loci, by and large. I think that it is likely that the brown-skinned Siberians did lose some alleles at particular loci (that is, they were fixed for loss of function variants), so that for true blackness to reemerge there needs to be new mutations which gain the function back. And as you likely know, gain of function is far less likely than loss of function. But that only explains why Native Americans don't get very dark. As I imply above, loss of function isn't all that hard. That's why albinos can be found in most human societies, they're an extreme mutant, but the same principle seems to be operative on many of the skin lightening genes. So why didn't Native Americans get pink? I think the fact that Siberians and Inuit are relatively brown suggests that extreme depigmentation is not always entailed by life at high latitudes. As many workers have suggested groups like the Inuit consume marine animals who are heavily loaded with Vitamin D, the lack of which is one of the presumed selective pressures driving depigmentation. That being said, most Native American tribes did not live next to the sea. And yet the recent selection events for genes such as SLC24A5 and OCA2 strongly implies that European have become very depigmented very late in prehistory, perhaps almost into historical periods! Why? I have proposed (following many others, such as L. L. Cavalli-Sforza) that the switch to agriculture resulted in a shift in diet and nutritional intake which entailed greater endogenous Vitamin D production by necessity. But there's a problem with this model: forms of agriculture existed in the New World as well, and spread up (eventually) into what became the eastern United States. Granted, the latitude of much of this region is about where the Middle East is, but even then it seems that the natives were relatively swarthy. I discount the notion that agriculture was too recent when SLC24A5 might have had selection coefficents on the order of 0.10. Perhaps the people of the New World, at least in North America, kept a more diverse diet, supplementing their agriculture with hunting and fishing to a far greater degree than in the Old World? Additionally, one might suppose that maize was nutritionally superior staple to wheat, barley, millet or rice (I have read that this is so). Ultimately these sorts of questions need to be addressed by a survey of the archaeological literature, as well as assessing the nutritional differences. I'll get to that at some point. But there's one last thing I thought of: disease. I can't really explain with SLC24A5 goes so far south in India. You see frequencies as high as 25% in Tamil Nadu. Vitamin D deficiency? Certainly nutritional stress is a major issue, but, one thing is for sure, South Asia is subject to a lot of disease in comparison to any other densely populated part of the world. Of the Old World civilizational hearths India was certainly the one weighed down by the greatest endemic pathogen load, in large part because it was so far south and so wet. So perhaps it was disease. Which brings us to Native Americans. Despite the recent uproar over syphilis, the New World was relatively pathogen free for humans. Granted, with greater population densities disease would have been a major issue among the agriculture populations of the New World, but there were structural reasons why they would have been less prone to epidemic outbreaks than Old World civilizations. The relative lack of domestic animals, the non-existence of closely related species (think of ape strains of viruses), the smaller and more fragmented population networks, and of course the fact that the original migrants probably only brought a small subset of the diseases of the Old World originally. Empirically we know that the Native Americans died like flies when the Eurasians showed up. Their civilization simply didn't prepare them for Old World plagues. What I'm proposing here is that disease was a major driver of skin color evolution over the last 10,000 years. Or, at least, the same loci which control and modulate melanin production are critical in immune defenses. I need to do a lot more digging for this to be anything more than a guess. But the disease angle seemed to be the last best hope in explaining why the New World was different. If they were subject to the same nutritional stress, why didn't they go down the same path as Eurasians? The reason may be that the path was being forged by the threat of disease (Vitamin D deficiency increases susceptibility to infectious agents), which was a less important parameter in the New World. Implausible as it may sound, it seems the most plausible of the various explanations to me. Note: If you are really curious about the topic, check out the many posts on skin color on this GNXP and the other. 1 - Even if Clovis First is debunked, it seems more and more likely that there are problems with genetic studies which claim that the earliest migrations date to 20-40 thousand years BP. 2 - All things being equal the rate of adaptive evolution is proportional to the extant genetic variation. If there is no genetic variation evolution has no raw material to work with. Labels: Genetics, Pigmentation
Thursday, January 17, 2008
Secret Of Scottish Sheep Evolution Discovered:
...gene and one copy of the light gene are quite large and also have quite high reproductive success. Sheep with two copies of the dark gene are larger still, but have poor reproductive success. Sheep with two copies of the light gene are small, but still have quite high reproductive success. This means that the two types of dark sheep although indistinguishable visually, vary in Darwinian fitness. The paper will be "A Localised Negative Genetic Correlation Constrains Microevolution of Coat Colour in Wild Sheep" in Science (not on the site yet). I wish I had some numbers to put on this...because I want to know the reproductive fitness of heterozygote black sheep vs. white sheep (ergo, the question mark after "balancing selection"). In any case, I wonder how it might apply to human pigmentation genes & selection. Consider the KITLG allele which results in dominance effects for light skin but recessive effects for light hair, and was subject to recent selection in Europeans. Or the affect of an OCA2 allele on eye & skin color. Labels: Genetics, Pigmentation
Friday, January 11, 2008
Via Dienekes, Eye Color, Hair Color, Blood Type, and the Rhesus Factor: Exploring Possible Genetic Links to Sexual Orientation:
The present study sought to expand the limited evidence that sexual orientation is influenced by genetic factors. This was accomplished by seeking statistical differences between heterosexuals and homosexuals for four traits that are known to be genetically determined: eye color, natural hair color, blood type, and the Rhesus factor. Using a sample of over 7,000 U.S. and Canadian college students supplemented with additional homosexual subjects obtained through internet contacts, we found no significant differences between heterosexuals and homosexuals regarding eye color or hair color. In the case of blood type and the Rh factor, however, interesting patterns emerged. Heterosexual males and females exhibited statistically identical frequencies of the A blood type, while gay men exhibited a relatively low incidence and lesbians had a relatively high incidence (p < .05). In the case of the Rh factor, unusually high proportions of homosexuals of both sexes were Rh- when compared to heterosexuals (p < .06). The findings suggest that a connection may exist between sexual orientation and genes both on chromosome 9 (where blood type is determined) and on chromosome 1 (where the Rh factor is regulated). What do you think? Seems more plausible that the likelihood of homosexual orientation is partly conditional upon the other genetic factors or physiological parameters; rather then there being a common causal root. If, as some argue, homosexuality is due to a relatively recent pathogen then its relationship to particular blood groups may simply be a coincidence of varied immune response of the different ABO & Rh antigens. I would be curious as to the blood group status of the mothers of gay men and women; perhaps it is simply due to physiological conflict (this might be related to sibling order). Like IQ it seems highly likely that there's a biological component to the variation, but color me skeptical of any locus of large effect. (FYI, I'm blood group A & Rh+) Related: Number of biological older brothers predicts male homosexuality, The biology of homosexuality, He She didn't give you gay, did she?, Pinker on the gay gene, Gavrilets' models of homosexuality, Gay sheep, forbidden science? and The gay gene & other considerations. Labels: Genetics, Pigmentation
Thursday, December 13, 2007
A few weeks ago p-ter posted on the fact that a gene that is implicated in blondeness in humans, KITLG, has a binding partner, KIT, within a similar affect in horses. There's a new paper out which I blog about here that shows that KITLG has a major affect on pigmentation in stickleback fish as well as humans, specifically showing a a partially dominant skin-lightening effect in African Americans in an admixture study. So like OCA2 this is now plausibly a case where selection for skin color could have driven secondary changes in phenotype (hair color). This makes more evolutionary sense since blonde hair is considered to be recessive, and so at a great selective disadvantage at low frequencies. In contrast, if skin-lightening is partially dominant it will be strongly exposed to selection (I'm skeptical of the dominance, they admit that more work needs to be done, but additivity has the same, less marked, advantage over recessivity). Note that KITLG shows up in tests of selection for East Asians too. You can find details for KITLG in this paper, Signatures of Positive Selection in Genes Associated with Human Skin Pigmentation as Revealed from Analyses of Single Nucleotide Polymorphisms, and showed up in Localizing Recent Adaptive Evolution in the Human Genome too. Note that the most recent paper, cis-Regulatory Changes in Kit Ligand Expression and Parallel Evolution of Pigmentation in Sticklebacks and Humans, is open access.
Labels: Genetics, Pigmentation
Thursday, November 22, 2007
In a few recent posts I've referred to the fact that variation on the OCA2 locus can predict about 3/4 of the eye color variation in the European population. Specifically, OCA2 is probably the quasi-Mendelian locus which is the culprit behind the classical dominant/recessive pedigree inheritance patterns which geneticists have long noted. The genomic region has also been subjected to a recent selection event. Why?
One model posits that the selection is directly for blue eyes. For example, some sort of sexual selection where blue eyes are strongly preferred. There's a problem with any model which posits selection for blue eyes: at very low frequencies selection on recessive traits is weak. That is, if you have alleles responsible for blue eyes extant at a frequency of 10%, only 1% of the population will express blue eyes (assumes random mating and a tighter correlation between the alleles and the phenotype as well as perfect dominance/recessiveness, all violated, but gets the logic across). So only 1 out of 10 blue eye causing alleles can be subject to selection. A way to get around this issue is population substructure, imagine that you have small demes drifting in all directions. A deme which drifts to a high or fixed frequency of blue eyes can then allow selection to operate strongly upon the allele responsible for this trait. This also requires specific meta-population dynamics so as to prevent these high frequency demes from being swamped out by gene flow from low frequency demes. Frankly, I'm really skeptical that a continent wide Shifting Balance process can really explain the third longest haplotype in the European genome. But there's another model, a bit simpler: the gene responsible for blue eyes is being selected for a another reason. Blue eyes are simply a byproduct, and that other reason is additive in its phenotypic expression so that even single copy variants are subject to the power of selection. I would hazard to guess that the most boring explanation here would be skin color. I've offered below that OCA2 does track skin color variation, but I've been pretty vague about this. The data isn't always easy to find, so I've repackaged Table 5 from A Three-Single-Nucleotide Polymorphism Haplotype in Intron 1 of OCA2 Explains Most Human Eye-Color Variation. Please note that there is a typo in the table in the paper, they have the correct data in the text, so I went by that.
Obviously there are other genes at work in regards to skin color, there's some population substructure which is probably lurking in the data, and the association of the variants themselves with an eye color aren't perfect either. That being said, this isn't the only study which does note that OCA2 has not only localized affects, but some global affects as well. Labels: Genetics, Pigmentation
Tuesday, November 20, 2007
Peter Frost states:
I suspect there is some incipient sex-linkage, i.e., European women may be somewhat likelier to have non-brown eyes and non-black hair. If this sex-linkage is mediated by prenatal estrogenization there may also be some impact on personality and temperament. But I really don’t know, and unfortunately there are still more questions than answers.  I've read Peter's book, Fair Women, Dark men, and it is a great collection of data. Also, he has theorized that European color variation is a byproduct of selection selection. So I have been primed to look for a trend where women seem to express blondism or light eye color at higher frequencies. But I just haven't found anything like that. In fact, I've found data which goes in the other direction, that is, females have a higher frequency of brown eyes! But this really clinched it for me: I've read Peter's book, Fair Women, Dark men, and it is a great collection of data. Also, he has theorized that European color variation is a byproduct of selection selection. So I have been primed to look for a trend where women seem to express blondism or light eye color at higher frequencies. But I just haven't found anything like that. In fact, I've found data which goes in the other direction, that is, females have a higher frequency of brown eyes! But this really clinched it for me: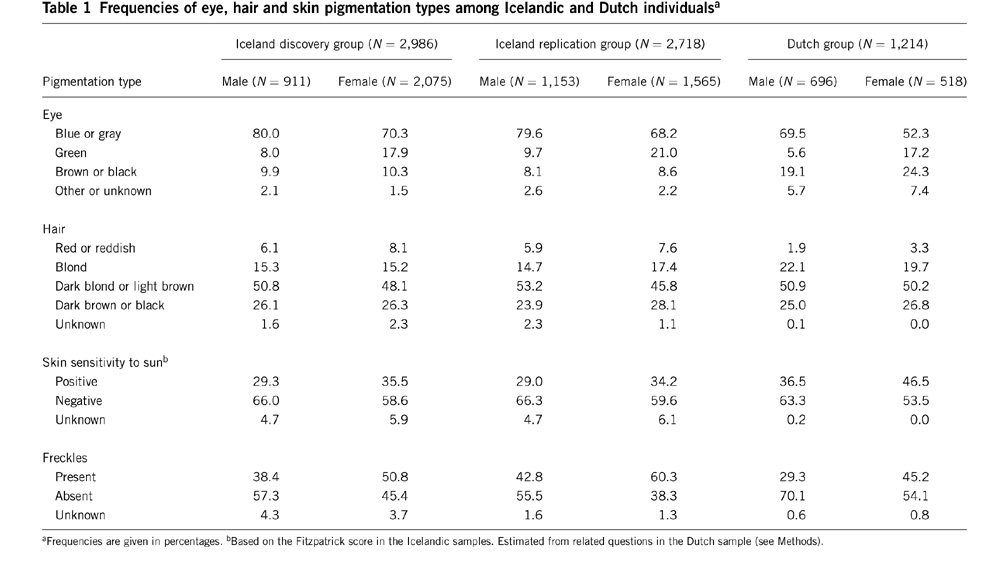 The source is this paper, Genetic determinants of hair, eye and skin pigmentation in Europeans. Note that women tend to score higher on skin sensitivity toward sun, which implies that they do have ligher skin. And as for hair color, well, perhaps there is a difference in how one judges blonde vs. brunette for males and females? I don't know. But the eye color data I've seen elsewhere and just dismissed it as small N or something like that. At this point my assumption is that there isn't really the sexual dimorphism in eye color that there is most definitely is in skin color. As for hair, I'm more open to this since it seems that it is subject to more genes, and there could be some hormonal factor as the tendency toward greater blondism in children and females is noted among Australian Aboriginals as well. Anyway, forget visual inspection. Here's the associations taking sex into account (from Table 4 of supplementary info): 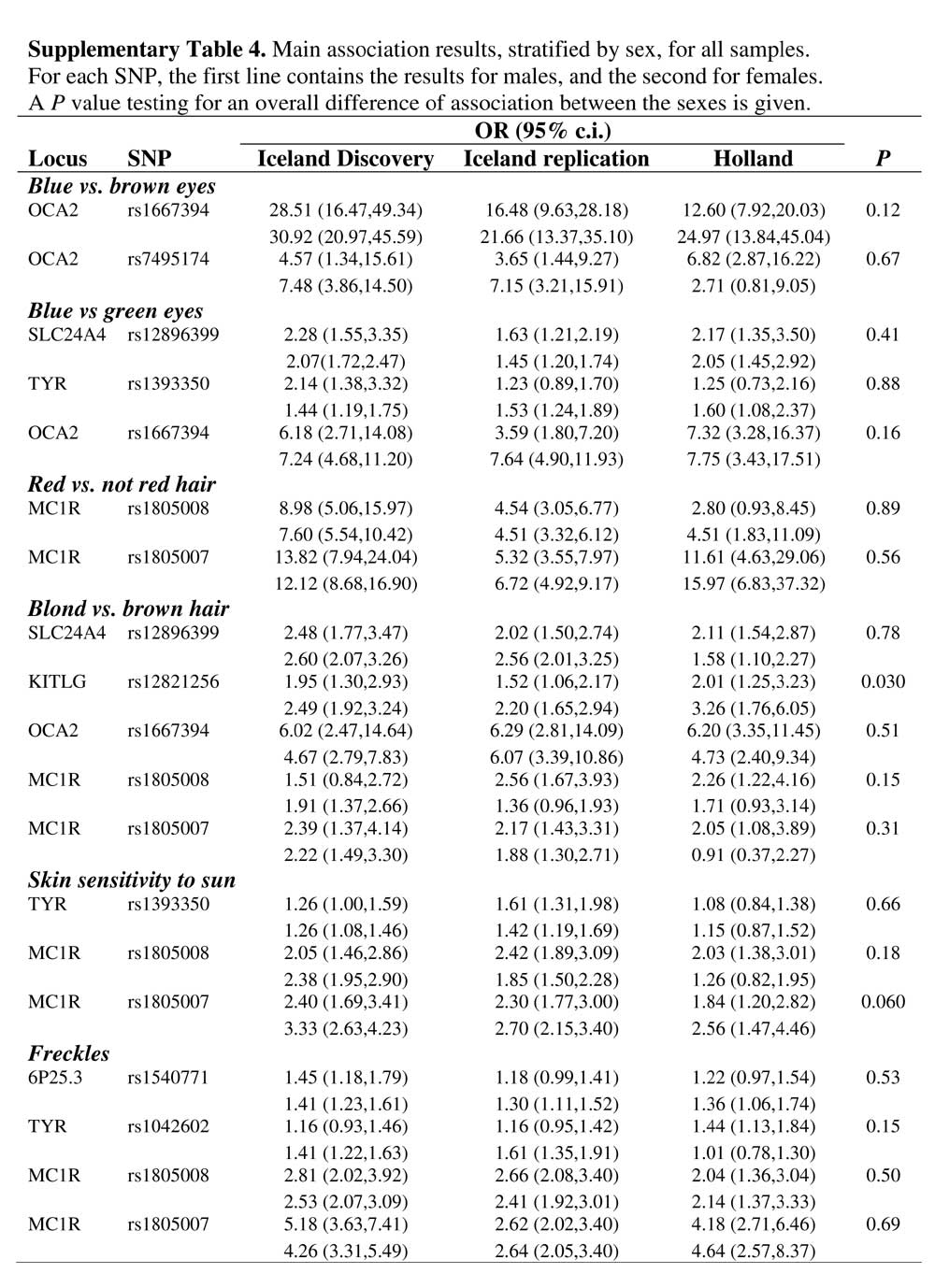 The authors don't want to make a judgment based on these data. But I'm not religious about 0.05 P values. And it looks like there's some action on KITLG anyhow. Labels: human biodiversity, Pigmentation
Monday, November 19, 2007
 I'm interested in blue eyes. Specifically, I wonder why they're around at all. Unlike blonde hair, there's only one region of the world where blue eyes are extant at high frequencies, and there is a pretty regular drop off as a function of distance. It seems that variants of OCA2 are associated with blue eyes in Europeans. If you check Haplotter it looks like the region around this gene has been subject to a powerful bout of recent selection (i.e., within the last 10,000 years). Why this selection? Well, there's no definitive explanation yet. But I don't want to focus on hypotheses for why OCA2 has been subject to selection as much as what correlates there seem to be with the phenotype of blue eyes. Specifically, behavioral correlates. I'm interested in blue eyes. Specifically, I wonder why they're around at all. Unlike blonde hair, there's only one region of the world where blue eyes are extant at high frequencies, and there is a pretty regular drop off as a function of distance. It seems that variants of OCA2 are associated with blue eyes in Europeans. If you check Haplotter it looks like the region around this gene has been subject to a powerful bout of recent selection (i.e., within the last 10,000 years). Why this selection? Well, there's no definitive explanation yet. But I don't want to focus on hypotheses for why OCA2 has been subject to selection as much as what correlates there seem to be with the phenotype of blue eyes. Specifically, behavioral correlates.There's a fair amount of recent work in this area, but reading Racial Adaptations I stumbled upon some older studies, and, a model to explain the outcomes which I'm not really qualified to judge. So I'm going to report and let you decide, and hopefully, inform (especially those of you with cognitive neuroscience backgrounds). So here I go. From page 66: ...eyes of different colors are related to differents in perception and innate behavior, as psychologists have discovered.23 Some of their tests have shown that dark-eyed persons are more aware of color and lighter-eyed ones of form; the former prefer bright hues and the latter lighter ones. Light eyes tend to envision panoramas, dark ones to concentrate on details I don't know anything about fashion or design, but I wonder if those of you who know this area (assman?) might be able to map some macrosocial trends back to these individual differences? After all, the vast majority of Scandinavians are blue eyed, and they've produced a fair amount of modern design. While Italians are well represented in the world of fashion and the arts. And the frequencies of eye colors are probably inverted in these two populations. In any case, moving on, later on the page: Behavioral variations are focused on the differences between "self-paced" and "reactive" responses to sudden stimuli. In the first subjects follow a well-known plan of animal behavior of pausing and deliberating before decision. In the second the subject flies into instant action (in animals), to attack or to flee. Of course, these behaviors are elaborated in man to govern many more-complex and subtler actions in speech and deed. That's a lot to throw at you, but pretty much line with more recent work. The author does note that these studies were performed upon subjects of European ancestry. Whatever differences one can see between groups of blue and brown eyed Europeans, obviously it wouldn't predict to other genetic backgrounds. East Asians tend to exhibit some of the same behavioral tendencies vis-a-vis Europeans that blue eyed Europeans exhibit vis-a-vis brown eyed ones. Obviously brown eyes can't explain this since East Asians have brown eyes. This isn't that strange, lots of the recent research in regards to human evolution suggests that East Asians and Europeans can converge upon the same phenotype via alternative genetic pathways. Blue eyes may simply be a byproduct of selection for another phenotype. But on to the author's model, which requires some knowledge of cognitive neuroscience and brain chemistry to evaluate. From page 74: Few people other than ophthalmologists seem to have looked at retinas, nor to have considered it remarkable that the fundus is of virtually the same color as the person's skin and for obvious reason that the underside of the retina is epidermis At this point the author draws upon some photographs of the retinas of various racial groups, and observes the variation in color. He takes lithographs of these photos and basically measures the amount of light which can penetrate them. Here is the exposition that is relevant: The Negro and the mulatto get 1.75 fcp; the Hindu and the American Indian 1.16 fcp; the brunet European 0.66 fcp; and the Chinese, the blond and the albino get 0.22 fcp. The Negro's and the mulatto's retinas let through eight times as much light as did those of the Chinese, the blond, and the albino. OK, here's the part where he lays out his argument for color and behavior: Once inside the cranial cavity, neural impulses produced by the visible light that has passed through the retinal screen follow one of two paths. One lot goes to the hypothalamus...This part of the brain is the primary control tower of the central nervous system for almost all of the self-starting and self-regulating activity of the body-the sleep cycle, body temperature, the digestive process, fighting and loving. That was a lot to throw out there, but the main reason I'm posting this is so that cognitive neuroscience people can throw cold water on this model or not. Obviously a lot has happened in neuroscience since the 1970s, when the author thought this idea up. Myself, I tend to get confused on the various biochemicals which modulate brain chemistry and all the different pathways and modulations, so clarification would be nice too. Two points I'd also like to point out. 1) the Chinese clustered with the blond European in terms of the retina light values. 2) If the amount of light which manages to get through the retina is a major issue, that can explain variation by latitude and climate in terms of temper and personality, since obviously the amount of sunlight and radiation which reaches the surface varies a great deal. Labels: human biodiversity, Pigmentation
In the interests of putting informative physical anthropological data on the web, here is something I scanned out of Racial Adaptations. The map below the fold combines both hair & eye color data.
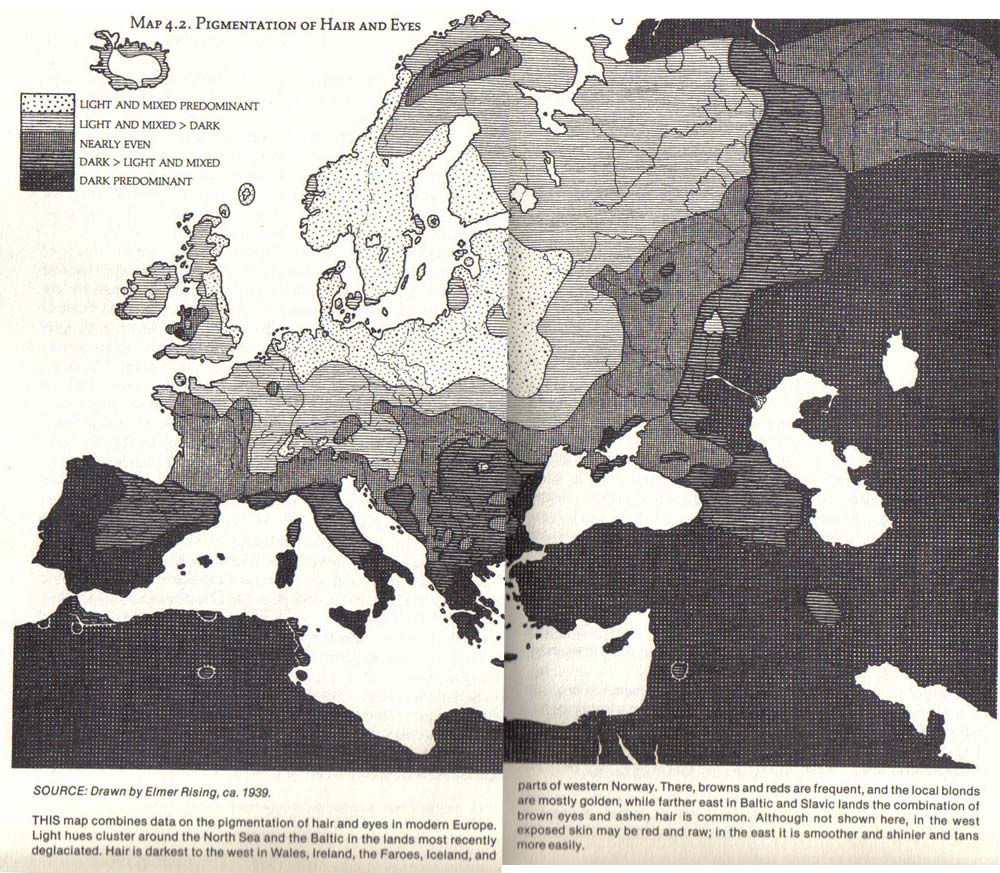 Labels: human biodiversity, Pigmentation
Sunday, November 18, 2007
 In PLoS Genetics, there's a report on the identification of a locus that leads to white coat color in horses. This locus is KIT, a proto-oncogene (ie. certain mutations in this gene lead to increased cell growth and sometimes cancer) important for the survival of melanoblasts early in development. It's a nice story on its own, and the authors have an interesting historical genetics perspective: In PLoS Genetics, there's a report on the identification of a locus that leads to white coat color in horses. This locus is KIT, a proto-oncogene (ie. certain mutations in this gene lead to increased cell growth and sometimes cancer) important for the survival of melanoblasts early in development. It's a nice story on its own, and the authors have an interesting historical genetics perspective:Two thousand years ago, the Romans already knew of the phenotypic differences of depigmented horses, which they described as candidus (white) or glaucus (grey). The Roman historian Tacitus described the use of sacred white horses for auguries by German tribes. The so-called white horse of the Saxons is depicted on the flags of the German states of Lower Saxony and North Rhine-Westphalia. It is thus of considerable historic interest to trace the origins of white horses, particularly because the nature of their white color can have different causes, some of which are KIT mutations such as those described here. We do not know whether the Roman terms candidus and glaucus actually correspond to our modern coat color designations of white and grey. Archaeogenetics on historic DNA samples may help to identify the genetic constitution of these horses.But something about that gene name bothered me: where had I heard it before? Ah yes, there was a recent report of an association with variants near KITLG (KIT ligand; ie. a binding partner of KIT) and blonde hair in humans. The genetic architecture underlying pigmentation isn't all that different across mammals, of course, so this isn't surprising. But still, it's nice to see these connections-- sometimes, biology makes sense. Labels: Genetics, Pigmentation
Wednesday, October 24, 2007
I've mentioned the emergence of the dog as a model organism in genetics-- the resources available now have made all sorts of questions easy (well, not easy, but comparatively easy) to answer. One such question: what makes certain breeds of dog black? The answer is now available online at Science: a small deletion in a gene called CDB103.
The gene is interesting because it encodes a novel ligand for MC1R, mutations in which cause red hair in humans (and lighter fur in mammoths). The melanocortin pathway seems to be polymorphic in all mammals studied, perhaps due to similar selective pressures on coat/hair color. The authors have a neat picture of mice they turned black by inserting copies of the variant allele, which raises the question: can any dog breed be turned black? And if so, how much will people pay for it? Labels: Genetics, Pigmentation
Friday, October 12, 2007
 The photo to the left is of a girl from Malaita in the Solomon Islands (Photo credit: Jonathan Friedlaender). The Solomons are in the heart of Melanesia, the "black islands." When I was a small child looking through anthropological books I would on occasion stumble upon photographs of very dark-skinned people from Oceania with light hair. Invariably they would not be light-brown Polynesians, but extremely dark Melanesians. You see references to this in C.S. Coon's old books as well. I've been digging for a few years into the nature of the trait of blondism in this part of the world, and I've come upon an interesting paper, Skin and hair pigmentation variation in Island Melanesia. Below the fold there is a map I've cut out of the skin & hair pigmentation. The photo to the left is of a girl from Malaita in the Solomon Islands (Photo credit: Jonathan Friedlaender). The Solomons are in the heart of Melanesia, the "black islands." When I was a small child looking through anthropological books I would on occasion stumble upon photographs of very dark-skinned people from Oceania with light hair. Invariably they would not be light-brown Polynesians, but extremely dark Melanesians. You see references to this in C.S. Coon's old books as well. I've been digging for a few years into the nature of the trait of blondism in this part of the world, and I've come upon an interesting paper, Skin and hair pigmentation variation in Island Melanesia. Below the fold there is a map I've cut out of the skin & hair pigmentation.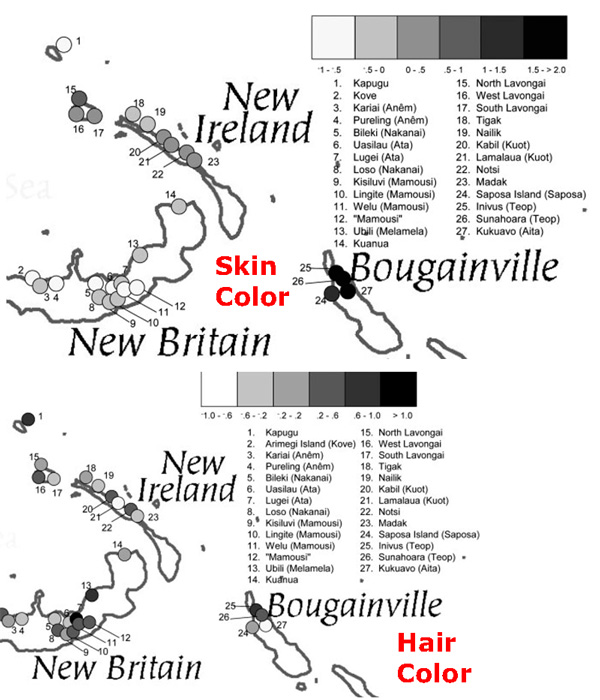 I'm intrigued by #27 on Bougainville, the Aita. From the text: An interesting hair phenotype that is sometimes seen in Island Melanesia (as well as among Australian Aborigines) is "blondism," in which individuals exhibit the characteristic darkly pigmented skin of the region while also having blond hair. This trait was most commonly observed in children whose hair generally darkened around puberty (Robins, 1991). However, in some cases, blondism persists into adulthood, although the hair appears somewhat darker than what is seen in children. The strikingly light hair of the Aita...is partially due to the high incidence of blondism among this group. So who are the Aita? They're an isolated "Papuan" group. In the context of Melanesia the appellation Papuan is simply a catchall for the host of ancient peoples who were extant in the region before the expansion of Austronesians within the last 5,000 years. Could the blondism be due to European admixture? From the text: The Aita live in an isolated mountainous region of northern Bougainville, and although the Ane'm had recently moved down to the coast they were originally an inland rather than shore-based group. It may be that the resistance of coastal groups to intermarriage with supposedly "darker" populations such as the Aita or Ane'm is really reflective of resistance to intermarriage with inland populations that are generally considered less sophisticated. Whatever the reason, this resistance may help strengthen the genetic differences observed between coastal and inland groups. The median skin color on Bougainville is amongst the darkest in the world, and the Aita are reputed to be among the darker residents of Bougainville. Additionally there is no report of blue eyes amongst these Melanesian peoples, which in Europeans is more common than blonde hair. Two papers, Melanesian mtDNA Complexity and Unexpected NRY Chromosome Variation in Northern Island Melanesia, shed some light on the genetic relationship of the Aita to the other groups in the region. They are it seems an ancient and rather isolated people who are geneticallyhomogeneous in a region characterized by a great deal of between group genetic variation. I am willing to hazard that presence of blondism in Melanesia is a function of the diversity of the region and its relative insulation from demographic sweeps which characterized much of the rest of the world. The Austronesians brought a more advanced culture and acculturated the coastal groups, but the ecology of Melanesia allowed the indigenous peoples, those termed Papuans, to maintain their existence inland. The high frequencies of an anomolous trait such as blondism among the Aita may simply be due to genetic drift through a population bottleneck. But a more curious question is the genetic architecture of blondism among these groups. The peoples of Bougainville are known to be functionally constrained in regards the genes which are implicated in controlling skin color variation. From Signatures of positive selection in genes associated with human skin pigmentation as revealed from analyses of single nucleotide polymorphisms: Our results showed that African populations tended to carry the ancestral alleles of the studied SNPs...and that human populations with dark skin colour tended to cluster together in the MDS and STRUCTURE analyses. The clustering of the Bougainville Islander population reflects the fact that this relationship is not (or at least not solely) caused by geography but also by the underlying pigmentation genes. Bougainville Islanders from Papua New Guinea are known to be one of the most highly pigmented people in the world...In our results they appeared to be closer to Sub-Saharan Africans populations - with whom they share the dark skin colour phenotype - than to the second Papuan New Guinea sample in the dataset, with whom they share their recent population history, as observed in several datasets based on neutral genetic variation from autosomal, Y-chromosomal and mtDNA analyses.... So how are they blonde? The genes which seem to control variation in human skin color seem to have some effect on hair color. Blondism in Europeans is simply due to low levels of eumelanin (brown) and pheomelanin (reddish), and the darkening of skin naturally has a tendency to upregulate the production of melanin globally so as to darken hair. How can the peoples of Bougainville retain dark skin while selectively downregulating the production to their hair? Well, we know that the region around OCA2 is implicated in a quasi-Mendelian disproportionate reduction in the pigment level in the iris. One assumes that something similar is theoretically feasible for hair.... Labels: human biodiversity, Pigmentation
Sunday, September 30, 2007
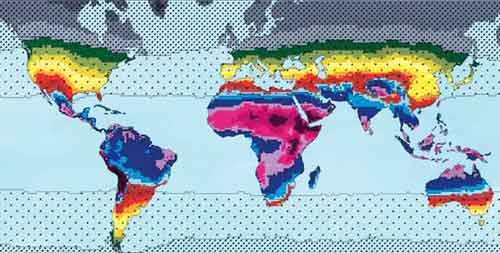 Update: I've added some geographic and ethnic notations to the ones that are relevant. For example, the Indian groups which are the darkest for their latitude turn out to be a Dalit and Tribal sample. In contrast, the other groups are more socially diverse. In South Afica the Capetown sample consists of mixed-race Coloureds. I've also added geographic data for places like Ireland, since I know there are readers who might be able to confirm with local knowledge (or disconfirm). Update: I've added some geographic and ethnic notations to the ones that are relevant. For example, the Indian groups which are the darkest for their latitude turn out to be a Dalit and Tribal sample. In contrast, the other groups are more socially diverse. In South Afica the Capetown sample consists of mixed-race Coloureds. I've also added geographic data for places like Ireland, since I know there are readers who might be able to confirm with local knowledge (or disconfirm).End Update From The Evolution of Skin Coloration by Nina G. Jablonski Figure 1: "The potential for synthesis of previtamin D3 in lightly pigmented human skin computed from annual average UVMED. The highest annual values for UVMED are shown in light violet, with incrementally lower values in dark violet, then in light to dark shades of blue, orange, green and gray...In the tropics, the zone of adequate UV radiation throughout the year (Zone 1) is delimited by bold black lines. Light stippling indicates Zone 2, in which there is not sufficient UV radiation during at least one month of the year to produce previtamin D3 in human skin. Zone 3, in which there is not sufficient UV radiation for previtamin D3 synthesis on average for the whole year, is indicated by heavy stippling." Below the fold I've reproduced a table that compares expected skin color and observed skin color for indigenous people. The expected is derived from a prediction equation which uses the observed values and combines them with the values from the UV map above: Predicted skin color = (annual average UVMED) X (-0.1088) + 72.7483 I also added a column which measures the difference between expected and observed and ordered it from populations which were lighter than expected to those which were darker than expected. Many of the values seem explicable via historical information (go to the paper and in the appendix you see what populations they used, that's important information); nevertheless, I am wondering about possibilities of different diet and its affect on skin color (more later)....
Notes: I'm skeptical of the accuracy of some of the reflectance measures. The authors report which ethnic groups they used for sampling in the appendices, so I would ask readers to look in there if they think some of these measures are questionable (I'll have a follow up post on this). They also assume that these "indigenous" peoples (which is, admittedly, a flexible definition) are well adapted to their local UV regime, and that other factors are controlled. Jablonski's thesis is that skin color is driven by two opposing forces: adaptation to high levels of UV which break down folate and increase birth defects, and, the need to synthesize vitamin D through the interaction of UV and biochemicals in the skin. Variation in diet and other possible selective forces aren't of much concern to her, and so she generated her expected skin color values assume that UV is the primary independent variable. My own hunch is that the far lighter than expected skin color across much of Asia is due to Vitamin D deficiency induced by the extreme carbohydrate biased diets of these populations. At this point this is just a tentative hypothesis, but, there has been selection for alleles known to be implicated in generating lighter skin in both South and East Asia within the last 10,000 years. Labels: Genetics, Pigmentation
Thursday, September 27, 2007
Sadly, but unsurprisingly, the little blonde girl photographed in Morocco turns out not to be missing British girl Madeleine McCann, but the daughter of a Berber farming family, who are said to have three other blonde children.
Most of us will have been vaguely aware that blonde hair and fair skin are not uncommon among the Berbers, but it has evidently come as a surprise to the general public. It is usually explained by a hypothetical element of European ancestry, whether from Roman slaves (as in this Daily Mail article), the Vandals, or more prosaically from the colonial occupation by French soldiers and government officials (who presumably didn't just twiddle their thumbs). I wonder if there is any hard genetic evidence? Y chromosomes might at least show whether the paternal ancestry is recently European. An alternative, and more interesting, explanation would be that the Berbers are the remnants of an older, more 'Caucasian', North African population. Labels: Pigmentation
Sunday, April 22, 2007
We previously reported that a measure of school achievement built from national test scores has a nearly perfect correlation with national IQ (at least in the range of scores tested). Subsequently, Lynn et al. (in press) published a very similar analysis:
This paper examines the relationship of the national IQs reported by Lynn & Vanhanen (2002, 2006) to national achievement in mathematics and science among 8th graders in 67 countries. The correlation between the two is 0.92 and is interpreted as establishing the validity of the national IQs. The correlation is so high that national IQs and educational achievement appear to be measures of the same construct. National differences in educational achievement are greater than differences in IQ, suggesting an amplifier effect such that national differences in IQs amplify differences in educational achievement. Controlling for national differences in IQ, slight inverse relationships of educational achievement are observed with political freedom, subjective well-being, income inequality, and GDP. However, public expenditure on education (as % of GDP) was not a significant predictor of differences in educational achievement. The IQ's Corner blog has an interesting note about forthcoming commentary. On a related note, recall that Templer & Arikawa (2006) reported a near perfect environmental correlation between national skin color and national IQ for old-world countries. An unfortunately confused commentary by Hunt & Sternberg accompanied the publication. They wrote: "We argue that the report by Templer and Arikawa contains misleading conclusions and is based upon faulty collection and analysis of data. The report fails to hold up for quality of data, statistical analysis, and the logic of science." The criticisms by Hunt & Sternberg are based largely on a misreading of Templer & Arikawa's methods, particularly the method for deriving national skin color values. A paper published in 2000 by Jablonski & Chaplin ("The evolution of human skin coloration") can more directly address these criticisms. Jablonski & Chaplin published a table of skin color reflectance values from many old world populations (Table 6, also see the appendix). I very crudely averaged values from the same country to make a new measure of national skin color. This measure of national skin color correlates with the skin color index of Templer & Arikawa at r=-.91 (the negative is not important here). The reflectance measure of skin color correlates with national IQ at r=.87. The school achievement measure of Lynn et al. correlates r=-.79 with the skin color index of Templer & Arikawa and r=.75 with the skin color reflectance values crudely averaged from Jablonski & Chaplin. Thus, the skin color values derived by Templer & Arikawa are well validated by an external data source and the national IQ-skin color relationship is found to be robust across two measures of national IQ and two measures of national skin color. Note that there are substantially more missing values in the school achievement and skin reflectance data sets (no imputation of missing values) with missing values skewed towards lower values of national IQ/school achievement and darker skin colors. Also note that the blind averaging use on the skin reflectance data most likely attenuates the correlations. Templer & Arikawa had two abstracts at the 2006 ISIR conference, which provide additional support for the validity of the measures and their relationships: source Correlations of Skin Color and Continent with IQ Donald I. Templer & Hiroko Arikawa The present study determined (1) the correlations between skin color and IQ across the countries of three different continents; and (2) the correlations of both skin color and continent in the three pair combinations with the three continents. The product-moment correlations between IQ and skin color were -.86 across the 48 African countries, -.55 across the 48 Asian countries, and -.63 across the European countries. When the 96 countries of Africa and Asia were combined skin color correlated -.86 and continent correlated .75 with IQ. The respective correlations were -.97 and .89 across the 81 countries of Asia and Europe, and -.71 and .54 across the 81 countries of Europe and Asia. In multiple regression continent yielded minimal increment to skin color in predicting IQ. In an earlier study (Templer & Arikawa, 2006a) skin color correlated more highly with IQ than racial category, but racial category yielded greater increments in multiple regression than did continent in the present study. The present findings, combined with previous research relating skin color and IQ (Templer &amp;amp;amp;amp;amp; Arikawa, 2006a; 2006b), indicate that skin color is a robust correlate of IQ in an international perspective. Empirical Support for Rushton's K Differential Theory Donald I. Templer & Hiroko Arikawa The purpose of the present study was to empirically substantiate Rushton's Differential K Theory that purports that groups of persons with K (in contrast to r) characteristics have a life history and reproductive strategy that includes higher intelligence, less reproduction, less sexual activity, better care of offspring, lower birth rates, greater life expectancy, better impulse control, and greater social organization. The present research intercorrelated national mean IQ, infant mortality, HIV/AIDS rates, birth rates, prevalence rates, and life expectancy in 129 countries in Africa, Asia and Europe. All of the correlations were substantial and in the expected direction. Also supportive of Rushton's theory is that there was only one factor which accounted for 75% of the variance and was labeled "K-r continuum." All five variables were correlated with an economic variable (per capita income) and a biological variable (skin color, which correlated highly with intelligence in previous research). Skin color correlated more highly with all five variables than per capita income so as to support the contention of Rushton that this continuum is biologically based. Factor analysis with all seven variables yielded one factor that accounted for 73% of the variance. Jason Malloy adds: Templer & Arikawa's research follows Lynn and Rushton in arguing that cold temperatures were a significant force in the evolution of human race differences in intelligence. I have stated some problems I find with this hypothesis here, although it is also largely consistent with the geographic distribution of global populations by IQ. A recent analysis by blogger Audacious Epigone adds yet another revealing data point to this association. Latitude (and hence colder climate) is associated with IQ not only cross-nationally (.67) but within the US as well. AE found a correlation of .70 between his measure of state IQ and the latitude of the most populous city in each of the 50 states. Furthermore intelligence is associated with latitude equally for both US whites and blacks (.52 and .51). It's not immediately apparent if and how this association is genetic or environmental. Either way it seems fair to seriously consider that global warming will provide yet another detrimental negative pressure on the intelligence of human populations in the coming decades. Labels: general intelligence, national IQ, Pigmentation, skin color
Friday, March 30, 2007
Some new data to throw into the argument about the origin of light skin (it seems that dark skin obviously arose when we lost our fur, seeing as functional constraint is strong in dark-skinned populations and unexposed skin in our nearest primate relatives is pink). From Dienekes:
Women have lighter skin than men do across a wide range of populations, even on the unexposed skin of the upper inner arm, possibly because of sexual selection by men for lighter-skinned women. If this hypothesis is true, human skin color should become more sexually dimorphic with increasing distance from the equator, since sexual selection for lighter skin in women would be less constrained by natural selection for darker skin in both sexes. Yet when Madrigal and Kelly (2006) analyzed skin reflectance data from 53 different samples, they found that the most dimorphic human populations were actually those of medium skin color at medium latitudes. Dienekes presents some values, and suggests that "In these data points it looks to me that Iranians and Kurds have the highest sexual dimorphism." I don't know what to make of that. Recall that sexual dimorphism often arises rather slowly as a genetically coded trait because obviously if you select for one sex at one end of the population range (sexual selection usually operates via male reproductive skew in most populations), their opposite sex offspring should also skew in that direction. A more complex genetic architecture which takes into account modifier genes on sex chromosomes (which will differ across sexes), or modulation via expression of sex hormones (which are dependent upon loci on those sex chromosomes ultimately, like SRY), seems necessary. Also, I was skimming through Nina Jablonski's Skin:A Natural History and on page 89 she states: "...one consistent observation is that women have lighter skin color than men. This is true for all indigenous peoples, even for those who are very dark-skinned, among whom such differences are not readily visible...." Jablonski's own hypothesis is presented in her paper The Evolution of Human Skin Color: ...the lighter skin pigmentation of females is needed to permit relatively greater UV light penetration of the integument for previtamin D3 synthesis. The extra calcium needs of females during pregnancy and lactation are met by increasing plasma concentrations of 1.25-dihydroxyvitamin D, which in turn enhances calcium absorption in the intestine.... Pregancy and lactation are critical periods which determine fitness. By focusing on this Jablonski gets a pretty good yield in terms of differential fecundity. She does not dismiss the importance of sexual selection as a secondary or subsequent factor in generating or heightening dimorphism. For the general interpopulation variation in skin color Jablonski focuses upon the balance between the need to prevent the breakdown of folate (due to UV) and produce enough vitamin D (also due to UV). She points to the Inuit as exceptions that prove the rule, insofar as their dark-skin is comprehensible because their diet is rich in vitamin D. From the genomic perspective we know that the architecture varies by location for similar phenotypic outcomes in regards to skin color. Even if the locus where a derived allele emerges is the same across two populations to generate the same phenotype (or contribute to the overall effect), that allele is often different, suggesting an independent mutational event. I would not be surprised if varied selective forces end up shaping human skin color variation. Though the correlation between UV and skin color is pretty clear, that may simply be the first principle component, with other factors rounding out the variation.... Update: Just an additional thought: a lot of the genomic data suggests recent selective sweeps on some of the genes for light skin (e.g., a variation of MC1R in China, the SLC45A2 derived allele in Europe, etc.). I think this is a big weakness in the model proposed by Jablonski, after all, it isn't like humans just showed up in northern Eurasia within the last 10,000 years. So what gives? I suspect that the lack of variety in the diets of agricultural peoples is an important factor. In other words, the more varied diet of hunter-gatherers (or late Ice Age big game hunters) didn't necessitate skin lightening to increase Vitamin D synthesis. Or, there are other selective pressures which we don't know about. Labels: Evolution, Genetics, Pigmentation
Wednesday, March 21, 2007
Over at her website Judith Rich Harris has posted her article, Parental Selection: A Third Selection Process in the Evolution of Human Hairlessness and Skin Color. When I asked Judy 10 questions I expressed some skepticism about this theory:
4) In your 2005 response to the Edge Question, "What Do You Believe Is True Even Though You Cannot Prove It?," you alluded to two things, 1) selection for light skin 2) hairlessness by parents in infants. When you pointed to these facts, did you do so in light of recent genetic work which suggests that dark skin might have evolved in humans as a response to loss of body hair? In other words, one trait would never been selected for if not for the other. Here is the abstract from the artice: It is proposed that human hairlessness, and the pale skin seen in modern Europeans and Asians, are not the results of Darwinian selection; these attributes provide no survival benefits. They are instead the results of sexual selection combined with a third, previously unrecognized, process: parental selection. The use of infanticide as a method of birth control in premodern societies gave parents - in particular, mothers - the power to exert an influence on the course of human evolution by deciding whether to keep or abandon a newborn infant. If such a decision was made before the infant was born, it could be overturned in the positive direction if the infant was particularly beautiful - that is, if the infant conformed to the standards of beauty prescribed by the mother's culture. It could be overturned in the negative direction if the infant failed to meet those standards. Thus, human hairlessness and pale skin could have resulted in part from cultural preferences expressed as decisions made by women immediately after childbirth. First, on a pedantic note, let me lodge my general protest in regards to the assumed decomposition of natural, sexual, and, Judy's putative third factor, parental selection. I'm reading some cognitive psychological work now about categorization and one thing that struck me as very apt in regards to how humans conceptualize the world is that we're always trying to break nature apart its joints, so to speak. Most regular readers know I'm a fan of consilience, and so I must reiterate and be a nag about the fact that many dynamic selective processes may bound in nature, but fundamentally they're all of a piece. As J.B.S. Haldane said: fitness is a bugger, naming selection is easy, characterizing it both accurately and precisely can often be very hard. The gene is the unit of selection, the various levels and dimension are all equal under God's eye. In any case, to the thrust of the hypothesis. First, in the past few years a lot of work has been done on skin color. Last year I observed that Armand Leroi's afterward to Mutants where he notes that we don't understand skin color will need a coda in future editions. Here's what we know, so far.... 1) In the 1960s human geneticists using classical pedigree analysis determined that 4-5 loci, genes, explained most of the intergroup variation between blacks and whites in regards to skin color. 2) In the past few years genomics has generally confirmed this view, there are a few loci of large effect (e.g., SLC24A5 in Europeans vs. non-Europeans) which explain intergroup differences in complexion. 3) But, equivalent phenotypic values can be attained via alternative genetic architectures, and this seems to have happened. In other words, light-skinned northeast Asians are not necessarily light for the same reason that Europeans are. Even if the change occurs upon the same locus, the allele or haplotype may be different. 4) Different evolutionary dynamics might affect the various loci. For example, in Europeans MC1R is highly polymorphic, suggesting either diversifying selection (e.g., frequency dependence) or a deep coalescence time (perhaps MC1R built up a great deal of variation during the residence of hominid lineages in Europe and modern humans simply assimilated the local depigmentation alleles?). 5) Also, some of these loci under selection seem to be relatively recent (e.g., perhaps within the last 10,000 years). Like LCT (lactase persistence) they leave a powerful imprint on the genome via a selective sweep. How does this square with the hypothesis presented above? First, some prelims. Selection of all sorts can be hard to get a grip on. After all, we bandy about selection coefficients, s, in a vacuum, when they vary within their environmental contexts. Environment in this case can mean the natural world, the social world and other genetic parameters (via genetic interactions). Negative frequency dependence also throws a monkey wrench into these processes by making the selection coefficient a function of the frequency of the traits. But we need to start from simple models and build up in complexity. The example of the !Kung woman who did not want to kill her light-skinned daughter is illustrative of Judy's hypothesis, but, it is simply a starting point. Nevertheless, I think it highlights a weakness: the genomic data is shedding light on the possibility that selection for loci which cause light skin (or, more properly explain a proportion of the intergroup variance) occurred long after the first humans settled the temperate zone. If the parental preference for light skin (which derives from the deep seated sensory bias which is also the root of sexual selection) existed prior to the arrival in the northern latitudes why is it that Eurasian populations seem to exhibit pulses of selection relatively late in history? One could make the argument, assuming that parental and sexual selection were paramount, that child and mate choice were simply not operative prior to this time period. Sexual selection works ideally through polygynous mating systems where there is a great deal of reproductive skew. Peter Frost has argued that blondeness in European females emerged through a form of sexual selection where males selected from a finite sample space of females because of the nature of the low latitude tundra, but the operative principle is the same, selection upon heritable variation. Perhaps within the last 10,000 years large boom and bust cycles in populations through the World Island has resulted in truncation selection events which reshaped the genomes (and generated high selection coefficients who show up in the long haplotypes)? I really don't know, but, I think one must say that it is more complex and contingent than a simple relaxation of functional constraint and the operation of innate preferences. Additionally, I don't see much exposition of the details of sexual selection theory in Judy's article. For example, runaway sexual selection occurs very fast, and is often quickly checked by functional constraints. I believe that her hypothesis about light skinned preference being very deep implies some sort of sensory bias. The intersexual difference in coloration is obviously real. There are biological reasons for this (hormone levels), but the cultural amplifiers of this tendency are rather universal. On this point the hypothesis is on strong ground, civilizations throughout the world seem to value female (relative) paleness. But, I think this point still goes back to my previous issue in regards to time scale: is this a phenomenon of the mass societies? In any case, I'll leave it at that. There's a lot to chew on here. My main point of contention with this hypothesis is that I think the time depth is off. The most current results out of genomics suggest repeated and independent evolutionary sweeps in northern Eurasia at various times generating the phenotype of light skin. If the preference for light skin is deep within our natures, even predating bipedalism, it seems that it should have manifested immediately with the move of H. sapiens sapiens to the northern latitudes 30,000 years ago. Labels: Evolution, Genetics, Human Evolution, Pigmentation
Thursday, February 01, 2007
Shelley Batts links to an interesting study looking at skin color and wages among recent immigrants to the US. On a scale of skin color, a single point lighter was good for about a 1% increase in wages, about equivalent to the effect seen for an additional inch of height.
The statistical analysis is pretty standard; nothing atrocious pops out at me (the obvious parameters-- country of origin, ability to speak English, occupation, etc-- were all included in the model), though they didn't control for IQ. But let's assume the effect is real. The hypothesis generally mooted for the association between height or attractiveness and wages passes through some sort of personality effect-- tall people are more confident, or more outgoing, or something along those lines. Is it possible there's a similar thing going on here? Labels: Economics, Pigmentation
Thursday, January 25, 2007
Blondes are not sexier: What the theory predicts and the data say
posted by
agnostic @ 1/25/2007 11:42:00 PM
Steve has an interesting post on assortative mating in which he purports, in passing, that blondes have greater sex appeal, citing Peter Frost's hypothesis that blonde hair was sexually selected in Northern Europeans (I'll post on the assortative theme later). A danger in discussing which traits might be sexually selected is that the ponderer will likely go with what they personally find sexy and ask why such things might be sexually selected, rather than work from an independent angle. For example, I am very picky about the upper eyelids -- if they have that half-moon shape, that really does it for me. I've heard Michael say once that he likes this feature too -- but then, we're probably weirdos, or at least that's the conclusion until someone can show that a large fraction of guys prefer this feature, and that there's good reason to think it was sexually selected. Unlike half-moon eyes, blonde hair color receives lots of attention as a potentially sexually selected trait, but is a key prediction met -- are blondes sexier?
Below the fold, I briefly review some theory but mostly present data from all winners in three beauty contests, which indicate no overrepresentation of blondes. I conclude that hair color is of weak importance at best in accounting for sexiness, that the role of sexual selection in accounting for hair color variation is also weak at best, and that the perception that men are more likely to find blondes sexy is due to a passing fad for blondes during the decade from the mid-1970s to the mid-1980s. First, Steve notes: One of the most common storylines in movies is this: the blonde debutante gets engaged to the blonde fraternity president, but then she falls hard for the tall, dark, and handsome boy from the wrong side of the tracks. The idea again is that blonde hair is sexy in females, not males. But just as common as the above scenario is the guy whose feelings for his fair-haired maiden waver once he becomes enchanted by a woman with coal eyes and raven tresses. For instance, in The Hunchback of Notre Dame (see here starting at line 57), the once pure priest Frollo describes how, after watching the gypsy Esmeralda dance and sing, he became so bewitched that he could not stem the tide of lust rising within him and fell madly in love with her. Shakespeare's "Sonnet 147" expresses a similar predicament: the speaker's got it bad for a bad girl! Not the first, nor the last. Interesting though literature and film may be, let's get to the theory and data. Starting with the predictions from theory, the hypothesis that icy climes select for greater sex appeal is probably wrong. Gangestad & Buss (1993) showed that people in more pathogen-wracked areas emphasize "good looks" more strongly, and these are generally not icy areas. Think about the rest of the animal kingdom: where are the sexy, showy specimens with the most ornate song patterns? Same answer: mostly pathogen-infested areas, the tropics, etc. Consider the quintessential animal with exaggerated sexually selected traits -- the peacock -- whose rarer variant is native to Southeast Asia, and whose more common variant is native to the world's germ-cauldron (South Asia). Hamilton & Zuk's (1982) explanation was that these traits signaled better health to mates, no small feat in such areas. So, the prediction is that sexual selection will be very weak in Northern Europe (defined as the non-Mediterranean countries), where blondeness reaches substantial frequencies. But even if sexual selection were a strong pressure there, what independently motivated evidence is there that blondeness is sexy, so that males who are sexually selecting would choose it over brunette hair? Again, the only good guess people have made is that sexually selected traits signal lack of being parasitized. For hair, though, this has mostly to do with the texture, lustruousness, and so on, not color -- although I'm willing to be corrected if someone knows of studies showing that blonde hair is more likely than dark hair to thwart the entry of pathogens into the scalp area. In any event, the main problem remains: in general, pathogen pressure is relatively very low in areas where blondeness is prevalent. Turning now to the data, I recently posted about the list of who Maxim magazine ranked the hottest women for 2006, and there was no evidence of overrepresentation of blondes. The same is true for the other "lad mags" that you see in drug stores. Now I look at two other datasets that are probably more informative than who Maxim thought was hot in 2006: the winners of the Miss Universe and Miss USA beauty pageants. (The Miss America competition is not primarily a beauty pageant, as looks account for just 35% of the score). Such lists are preferable for testing the "sexy blonde" hypothesis since the individuals represent a very elite level of eminence. I looked up galleries of the winners, and if a girl's hair color wasn't clear from that, I did a Google image search for her. I judged overrepresentation based on the frequency of light hair according to Peter Frost's map at the Wikipedia entry for hair color. For Miss Universe (gallery), there are 56 data points: 12 (21.4%) have light hair, 43 (76.8%) have dark hair, and 1 (1.8%) is pretty in-between. Now, 21.4% is surely a greater fraction of blondes than there are worldwide, but remember that Miss Universe doesn't represent the entire world -- it's mostly Europe and its offshoots, plus the white and mestizo populations of Latin America, and a tiny handful of East Asian countries (not China). For the non-Mediterranean areas of Europe, 21.4% is on the low-end of normal, but on the high end of normal for the Mediterranean (and so, for the mostly Mediterranean-looking Latin Americans who compete). I interpret this as supporting the null hypothesis of no effect of hair color on sexiness. As for Miss USA (gallery), there are 60 data points: 17 (28.3%) have light hair, 38 (63.3%) have dark hair, and 5 (8.4%) have borderline hair. Although the fraction is larger here, remember the US is much blonder than the Mediterranean and Latin American countries who are also big contenders in the Miss Universe competition. Because the vast majority of the US population has been Northern European since the pageant began in 1952, we should determine overrepresentation based on the Northern European areas of Peter Frost's map. Doing so, we see that 28.3% is easily at expectation, and if anything is a bit on the low-end of normal for a predominantly Northern European population. Again, this result supports the null hypothesis. In sum, we note that when put to a stringent test, blondes appear no sexier or uglier when compared to brunettes. Datasets such as Miss Universe and Miss USA are particularly instructive since the bar is set rather high. Then whence the perception that men find blondes sexier? There is an interesting temporal wrinkle in the data -- blonde winners are not evenly distributed in either dataset. For Miss Universe, from 1952 - 1974, 17.4% of the 23 winners are blonde; from 1975 - 1984, 60% of the 10 winners are blonde; and from 1985 - Present, either 8.7% or 13.0% of the 23 winners are blonde (depending on whether you are generous and code the 1 borderline girl as blonde). There thus appears to be a general lack of interest in blondes (and if anything, a dispreference for them), punctuated by a decade where blondes were very fashionable. Does the same pattern show up in the Miss USA dataset? Pretty much. From 1952 - 1973, 25% of the 24 winners were blonde; from 1974 - 1986, 50% of the 14 winners were blonde; and from 1986 - Present, 18.2% of the 22 winners were blonde. We note again the spike in blonde fashionableness from the mid-1970s to the mid-1980s. I suggest that those who came of age during this Blonde Decade -- those who were born between roughly 1955 and 1970 -- may have unwittingly projected their perception of the sexiness of blondes onto time periods for which the view is not true. Combine this with the theoretical problems noted earlier, and it seems likely that sexual selection's role in increasing the frequency of blondeness is weak at best. That still doesn't answer the question of why blondeness evolved -- though I'll leave that for another post (or someone else can take it up). The explanation that I (and others) find most convincing for now is based on Jerome Kagan's work, starting in the mid-1980s, which has showed that light irises correlate with behavioral inhibition, suggesting that in Northern Europeans there was selection for different values of certain personality traits, which happened to also affect their eye & hair color. Appendix: Hair color data for Miss Universe and Miss USA winners Miss Universe (L = light, D = dark, M = borderline) L 1952-Armi Helena Kuusela Kovo-Finland D 1953-Christiane Magnani (Martel)-France D 1954-Miriam Jacqueline Stevenson-USA L 1955-Hillevi Rombin-Sweden D 1956-Carol Laverne Morris-USA D 1957-Gladys Zender Urbina-Peru D 1958-Luz Marina Zuluaga-Colombia D 1959-Akiko Kojima-Japan D 1960-Linda Jeanne Bement-USA L 1961-Marlene Schmidt-Germany D 1962-Norma Beatriz Nolan-Argentina D 1963-Ieda Maria Britto Vargas-Brazil D 1964-Kiriaki “Corinna” Tsopei-Greece D 1965-Apasra Hongsakula-Thailand L 1966-Margareta Arb Arvidsson-Sweden D 1967-Sylvia Louise Hitchcock-USA D 1968-Martha Maria Cordeiro Vasconcellos-Brazil D 1969-Gloria Maria Diaz Aspillera-Philippines D 1970-Marisol Malaret Contreras-Puerto Rico D 1971-Georgina Rizk-Lebanon D 1972-Kerry Anne Wells-Australia D 1973-Maria Margareta Moran Roxas-Philippines D 1974-Amparo Muñoz Quesada-Spain L 1975-Anne Marie Pohtamo-Finland D 1976-Rina Messinger-Israel D 1977-Janelle “Penny” Commissiong-Trinidad/Tobago L 1978-Margaret Gardiner-South Africa D 1979-Maritza Sayalero Fernández-Venezuela L 1980-Shawn Nichols Weatherly-USA L 1981-Irene Lailin Sáez Conde-Venezuela D 1982-Karen Dianne Baldwin-Canada L 1983-Lorraine Elizabeth Downes-New Zealand L 1984-Yvonne Ryding-Sweden D 1985-Deborah Carthy-Deu-Puerto Rico D 1986-Bárbara Palacios Teyde-Venezuela D 1987-Cecilia Carolina Bolocco Fonck-Chile D 1988-Porntip Nakhirunkanok-Thailand L 1989-Angela Visser-Holland D 1990-Mona Grudt-Norway D 1991-MarĂa Guadalupe “Lupita” Jones Garay-Mexico D 1992-Michelle McLean-Namibia D 1993-Dayanara Torres Delgado-Puerto Rico D 1994-Sushmita Sen-India D 1995-Chelsi Pearl Smith-USA M 1996-Yoseph Alicia Machado Fajardo-Venezuela D 1997-Brook Antoinette Mahealani Lee-USA D 1998-Wendy Rachelle Fitzwilliam-Trinidad/Tobago D 1999-Mpule Keneilwe Kwelagobe-Botswana D 2000-Lara Dutta-India D 2001-Denise Marie Quiñones August-Puerto Rico D 2002-Oksana Fyodorova (Oxana Fedorova)-Russia (dethroned) D ---Justine Lissette Pasek Patiño-Panama D 2003: Amelia Vega Polanco-Dominican Republic L 2004: Jennifer Hawkins-Australia D 2005: Natalie Glebova-Canada D 2006: Zuleyka Jerris Rivera Mendoza-Puerto Rico Miss USA D Jackie Loughery 1952 D Myrna Hansen 1953 D Miriam Stevenson 1954 L Carlene King Johnson 1955 D Carol Morris 1956 D Leona Cage 1957 L Charlotte Sheffield 1957 M Eurlyne Howell 1958 D Terry Lynn Huntingdon 1959 D Linda Bement 1960 D Sharon Brown 1961 D Macel Wilson 1962 L Marite Ozers 1963 L Bobbie Johnson 1964 L Sue Downey 1965 D Maria Remenyi 1966 D Sylvia Hitchcock 1967 D Cheryl Ann Patton 1967 D Dorothy Anstett 1968 L Wendy Dascomb 1969 D Debbie Shelton 1970 D Michele McDonald 1971 M Tanya Wilson 1972 D Amanda Jones 1973 L Karen Morrison 1974 D Summer Bartholomew 1975 D Barbara Peterson 1976 L Kimberly Tomes 1977 L Judi Andersen 1978 M Mary Therese Friel 1979 L Shawn Weatherly 1980 L Jineane Ford 1980 L Kim Seelbrede 1981 D Terri Utley 1982 D Julie Hayek 1983 D Mai Shanley 1984 D Laura Martinez-Herring 1985 L Christy Fichtner 1986 D Michelle Royer 1987 D Courtney Gibbs 1988 D Gretchen Polhemus 1989 D Carole Gist 1990 M Kelli McCarty 1991 L Shannon Marketic 1992 D Kenya Moore 1993 D Lu Parker 1994 D Chelsi Smith 1995 D Shanna Lynn Moakler 1995 D Ali Landry 1996 D Brook Lee 1997 D Brandi Sherwood 1997 M Shawnae Jebbia 1998 D Kimberly Ann Pressler 1999 D Lynnette Cole 2000 L Kandace Krueger 2001 D Shauntay Hinton 2002 D Susie Castillo 2003 L Shandi Finnessey 2004 D Chelsea Cooley 2005 L Tara Elizabeth Conner 2006 Labels: babes and hunks, Pigmentation
Friday, September 29, 2006
A question from Jason Malloy prompted a quick search of the GSS for data on the cause of the Black-White IQ gap. In 1982, the GSS characterized the skin color of Black participants on a 5-point scale (1:very dark brown to 5:very light brown). The very dark/light categories consist of only 50 and 14 individuals, respectively, and so in the following analysis I merged them with the dark/light brown categories, to give three COLOR levels: dark, medium, and light. In the web application, use COLOR(r:1-2;3;4-5) instead of COLOR. The WORDSUM variable is a 10 question vocabulary test, which I'm treating as a proxy for IQ. It is correlated with educational attainment (~.4), and also correlates (~.4-.5) with tests of reasoning and basic knowledge that were given in some years. These other tests are not available for 1982. In the all-subject all-year GSS data set, WORDSUM varies by SEX, and in 1982 COLOR also varies by SEX. Thus, SEX is controlled for in each analysis. WORDSUM is lower in the youngest and oldest age groups, so an AGE(25-65) filter was used.
Table 1. Mean WORDSUM score by COLOR and SEX with ANOVA
color indicates T-statistic, and thus p-value
We can quantify the effect size of each skin color class using Cohen's d statistic, which measures the mean difference in standard deviation units. In the 1982 dataset, the overall d for the Black-White gap on WORDSUM is -0.63 (among males d=-0.51, among d=-0.74). For comparison, the 1982 male-female gap among Whites is d=-.12, favoring females. Table 2. Effect size (d) of COLOR on WORDSUM using "light" as a control group
We can also use Whites as the control group. Table 3. Effect size (d) of COLOR on WORDSUM using Whites as a control group
Thus, there are substantial (moderate to large effect size) differences in WORDSUM scores between the darkest and lightest Blacks in 1982. As reported by Rushton and Jensen (2005), Shuey (1966) reviewed 18 studies which used skin color as a measure of racial admixture to compare with IQ. Of those 18, 16 found a significant effect of the kind found here, but the overall correlation with IQ was low (r=.1). In this data, the COLOR WORDSUM correlation is r=.31 among males and r=.18 among females, with an overall correlation of r=.23. Off the top of my head, I'm not certain what the expected correlation would be between IQ and skin color among Blacks for a given measure of "between-group heritability" (BGH) as described by Jensen (1998). I'll leave it as an exercise for our mathematically skilled commentators to derive a formula for this relationship and to evaluate the signficance of this finding in explaining the cause of the Black-White IQ gap. Labels: Economics, GSS, IQ, Pigmentation, race
Saturday, September 23, 2006
 ...here. Like the study Razib linked to a couple days ago, this one looks for signatures of selection in a number of genes suspected to play a role in the generation of natural human skin color variation. And also like the previous study, they find that different genes are implicated in derived light skin color of east asians and northern europeans (see the figure on the left for a crude representation of this). Labels: Genetics, Pigmentation
Thursday, September 21, 2006
 Identifying genes underlying skin pigmentation differences among human populations:
There are many posts about skin color in the GNXP archives. This should not surprise. The fact that SLC24A5 was positively selected for in Europeans and correlates with light skin, but remains ancestral in both Africans and East Asians, while the Arg163Gln variant of MC1R seems under selection in East Asians, but under relaxed selection or possibly diversifying selection in Europeans suggests that different genetic strategies are scaling the same phenotypic fitness landscape. Additionally, two different strategies suggests to me that gene flow has been low enough so that one best case strategy (i.e., the highest fitness peak in a large admixing population) isn't automatically selected across Eurasia across the common genetic background. You know what they say, selection is stochastic.... Labels: Pigmentation, Population genetics
Tuesday, August 01, 2006
Via Yann, The golden age of human pigmentation genetics, in Trends in Genetics. Below the fold you'll find the table which encapsulates most of the relevant information.
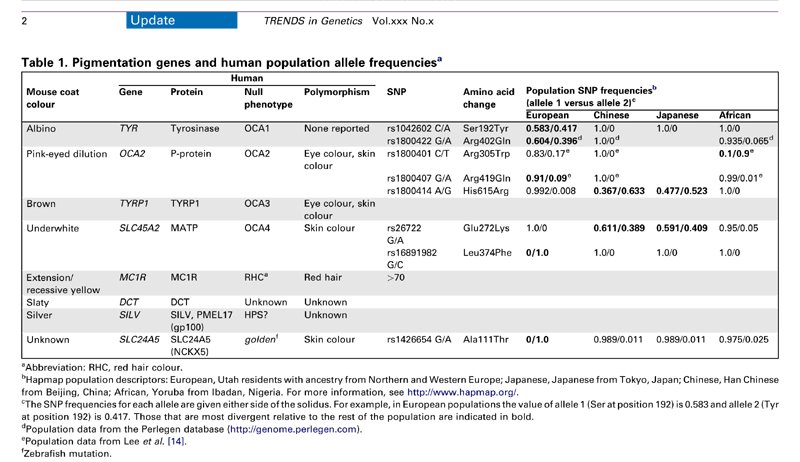 Labels: Genetics, Pigmentation
Friday, May 12, 2006
Dienekes points me to this piece in The American Journal of Physical Anthropology which concludes that there isn't greater sexual dimorphism in skin color in areas where social/sexual selection could presumably operate because of relaxation of natural selection. Remember that sexual dimorphism tends to evolve very slowly (you need to have sex-linked or developmentally modified loci since men and women share almost all the same genes [except for the Y]), perhaps an order of magnitude more slowly than standard phenotypic evolution given the same selection pressure.
Labels: Pigmentation, sex differences
Wednesday, May 10, 2006
The Finns and Swedes are arguing eye color. So, Eye colour: portals into pigmentation genes and ancestry:
One major locus, but lots of little modifier loci which an account for shades of green, gray, etc. (not to mention epigenetically derived variation). Also, remember this isn't MC1R, so that is why eye color and hair color are inherited independently. Take that Jaakkeli! You lemur-eyed freak! Labels: Genetics, Pigmentation
Saturday, April 22, 2006
In December I posted a hypothesis about skin color evolution in East Asians and Europeans in light of new research which suggested that Europeans were fixed for a particular locus which explained a great deal of the variation between themselves and Africans. The facts are like so:
1) There are probably around 5 loci of major effect that explain skin color variation in human beings (older pedigree analysis as well as new genomic data seems to be focusing on a number in this range). 2) One of the major loci, MC1R, is highly polymorphic in Europeans, constrained in dark-skinned populations to the ancestral consensus sequence (Africans, to a lesser extent in South Asians), and being driven toward fixation on a derived allele in East Asians. 3) A new locus in Europeans seems fixed and is responsible for the skin color depigmentation, but in both East Asians and Africans this locus is in an ancestral condition. From these data I hypothesized that East Asians and Europeans were genetically isolated so they reached the same fitness optimums (assuming light skin has selective benefits at high latitudes) via different genetic architectures. In Europeans MC1R is diversified (either through frequency dependent selection or relaxation) because SLC24A5 handles the phenotype. In East Asians SLC24A5 is in an ancestral state, so MC1R has been coopted via the derived allele to generate the same end via different means. I think I was wrong, and the evil Finns are to blame...they exhibit a rather high frequency of the MC1R allele which confers light skin in East Asians, but they're in Europe! The idea that I threw out there is really contingent on powerful genetic barriers, beneficial alleles have a way of sweeping everywhere that they confer fitness all things being equal. Lactose tolerance is spread throughout Eurasia, and not because of massive population movements, while various malarial resistance alleles are pretty common across regions just where malaria is endemic. Perhaps there was a massive ice sheet in the middle of Eurasia, and Finns are the byproduct of an admixture event, but my understanding is that MC1R has been under positive selection for around ~10,000 years in East Asians. That's pretty recent, and I don't think that the genetic barriers across Eurasia were that strong for that period, they should been able to "borrow" SLC24A5 instead of throwing up their own trick de novo. Other shit is going down...and I'm sure that pleiotropy has its dirty hand involved here. But I was wrong, though I did give it a good college try :) Labels: Genetics, Pigmentation
Thursday, April 20, 2006
Now, we know that Europeans seem to be light in skin color because of fixation of a derived allele at a locus other than MC1R. In contrast, East Asians are "ancestral" at the locus where Europeans are "derived," but they show evidence of strong positive selection for the Arg67Gln haplotype on MC1R. Here are Arg67Gln frequencies by population (source):
Inuit 92.5% Japan 76.7% ***Finland 30.0%*** Papua New Guinea 12.5% Saami 11.5% Sweden 7.7% Ireland 6.5% UK Asian Indians 4.5% England 3.1% Italy 2.9% Southern India 2.3% Ivory Coast 0.0% The Gambia 0.0% UK Africans 0.0% Sardinia 0.0% Along with data from TAT C it makes you wonder. They used to say you scratch a Russian and you find a Tatar, well, if you scratch a Finn.... Labels: Genetics, Pigmentation
In the GNXP Forum I have placed a PDF titled skincolorsexual.pdf. It is the full PDF of Sexual Selection as a cause of human skin color variation:
Since I've put the full paper online, I suggest people actually read it before they comment (*hint*). Labels: Evolution, Pigmentation
Saturday, March 18, 2006
I read Peter Frost's paper on blonde hair and sexual selection awhile ago. I don't have much to say on it that I didn't already say in the previous conversation, I think ascertainments or suppositions in regards to sexual selection in the evolution of the human species and our phenotype(s) will be a lot more fruitful when we have the priors in in regards to the organization of our genome and its deep time history locked down in more detail. I'm posting now because I did think Peter put a lot of interesting data out there, and I am reproduced two of the maps in his paper, which depict the frequency of blonde hair and blue eyes respectively in northwest Eurasia. Nothing surprising, but some concrete percentages for any future discussions on the topic. You'll find it below the fold.
The only substantive point I'd like to add is that Peter points out that the emergence of 30 polymorphisms extant within the European populations on MC1R as a consequence of release of functional constraint is implausible (the locus implicated in melanin regulation which in East Asians and other populations tends to be far less polymoprhic, or nearly monomoprhic, as in Africa). He contends that the current level of hair color diversity in Europe would have taken 850,000 years to develop if one assumes release of constraint was the primary factor, and of course H. sapiens sapiens has been in Europe no longer than 35,000-40,000 years (one assumes that release of constraint would not have occurred prior to leaving the lower latitudes). But, I would like to point out that H. sapiens neanderthalensis was resident in Europe for ~250,000 years. This isn't enough time for a neutral-only model to generate the diversity, but life isn't always an either or scenario, and one could imagine that factors other than in situ mutation or selection upon standing genetic variation might have played a role in the diversification (or perpetuation of diversity) of European color phenotypes. Update: Greg points out that Neandertal (H. heidelbergensis) precursors might have lived in Europe as early as 800,000 years B.P. So that is a point to consider. Also, I have to add this in the "scientists don't get evolution" files, I recall that one paleoanthropologist posited that Neandertals were dark-skinned because their skeletons seem to exhibit features similar to rickets. At the time (I was 16 when I read this) I wondered how plausible it was that 800,000 years of hominin residence in high latitudes wouldn't have selected against traits which predispose one to rickets. Obviously it isn't plausible.   Credit: Beals et al., An Introduction to Anthropology, 3rd ed. Labels: Pigmentation
Thursday, February 23, 2006
 Update: More comments here, here and here. Update: More comments here, here and here. End update Desidancer and Diana both pointed me to this story about a mixed race couple who gave birth to daughters of very different phenotypes. The explanation in the story is about right, the loci which give you a gestalt impression of racial identity are a tiny sample of your overall genome. In the story it is reported that 7 genes control skin color and F1 (first generation) hybrids should carry half of the variants of each race (since their parents contribute exactly 50% of the genes to each). But the F2 generation can come out as a range of combinations, so it stands to reason that mixed-race couples will have children who vary a great deal in phenotype as the alleles resegregate themselves into alternative combinations. I lay out the details in By the Punnett Square. South Asians, who often exhibit a wide range of color variation from near white to near black, should not be surprised at this sort of dynamic, as the variance within a family can be rather large in complexion. In any case, independent assortment implies that even if the coloration of these two children reflects one of their ancestral ethnic groups, other traits do not necessarily line up in such a fashion. Since the twins above are babies it is hard to discern facial traits (they just look like babies), but I wouldn't be surprised if the "white" and "black" baby had facial traits that were more equidistant to the metrics of whites and blacks (this would be expectation, though you expect a lot of variance still). There is a reason that in much of the southern part of the New World where admixture between Africans, Europeans and American indigenes is common, there are dozens of definitions for racial phenotypes, because the full range of appearance is expressed in a large enough mixing population (eg, there are terms for people with Negroid facial features and hair form and Nordic coloration and European facial features and hair form and Sub-Saharan African coloration). By the way, as personal stories like this become more common hopefully we'll stop hearing about how everyone in the future will be brown because of admixture. More values at each variable will result in an increase in variance for the distribution, not a decrease. Update: OK, I think I need to repost this link, The incidence of superfecundation and of double paternity in the general population:
In other words, double paternity is a possible explanation, but if the number of loci in question is seven or less than this is surely will the expected range due to variance emerging from heterozygosity in the parents1 (and we are only looking at skin color here from what I can tell, the two infants are still of the "baby race," other features are not at play). I don't think that the expectation of double paternity, evening adjusting for SES, approaches the probability that "white" and "black" color genes will resegregate in this fashion. Update II: David points out that this is being reported in the British newspapers, which does alter your Bayesian priors, but walk around Lowell, Massachusetts, and observe the range in phenotype of the Cape Verdian community. The general point still stands. 1 - Fisher's 1918 paper dealt in large part with the variance expected from heterozygous parents. Labels: Pigmentation, race
Saturday, January 28, 2006
Since very few of you have likely read Fair Women, Dark Men: the Forgotten Roots of Racial Prejudice by Peter Frost, I'd like to you point you to his website, where he introduces many of his ideas in a series of essays. Steve also has an essay on based on Frost's ideas, and you might find this paper by Frost, European hair and eye color A case of frequency-dependent sexual selection?, of interest.
Labels: Pigmentation, sex differences
Friday, December 16, 2005
A fascinating paper just came out in Science, SLC24A5, a Putative Cation Exchanger, Affects Pigmentation in Zebrafish and Humans. Heather L. Norton is one of the authors listed, so she knew very well what she was talking about when she suggested that there was far more to skin color variation than MC1R. Here is the eye popper from the text: "Based on the average pigmentation difference between European-Americans and African-Americans of about 30 melanin units (33), our results suggest that SLC24A5 explains between 25 and 38% of the European-African difference in skin melanin index." I have pointed to models before which suggest ~4-5 loci that control skin color, this work does nothing to falsify that, but, it does show that between populations the variation in coloration in humans might be due to alleles of large effect which are differentially fixed. Evaluated over the whole species it is also certainly true that various alleles account for a different percentages of the genotypic component of variation, with many small effect modifier alleles likely hovering in the background of populations which are fixed for alternative alleles of large effect. Another important point is that the authors point out that both Africans and East Asians exhibit the ancestral allelic state, while Europeans are derived (a mutant descendent form), so the implication is that light skin is generated in East Asians by alternative genetic conformations. I have already pointed out that East Asians seem to be under strong selection, and moving toward fixation for Arg163gln MC1R allele (in contrast, Europeans are highly polymorphic for this locus). This points to the reality that evolution, or precisely, selection, explores a large phenotype & genotype space with multiple fitness peaks, and the expected outcomes might be random in nature when the options are constrained to the various peaks. The title of the post comes from the author's observation that "This region [of the HapMap -R], which contains several additional SNPs with high-frequency differences between populations, was the largest contiguous autosomal region of low heterozygosity in the European (CEU) population sample...This pattern of variation is consistent with the occurrence of a selective sweep in this genomic region in a population ancestral to Europeans." The low heterozygosity seems to be due to powerful directional selection which dragged a large number of adjacent SNPs within that haplotype block to fixation.
Finally, I want to add that this is a cool paper partly because of its broad methodological scope. Instead of just scanning the HapMap the authors also confirmed the developmental genetic expression via an animal model, zebrafish, which seems to exhibit the same variation in coloration on this locus as our own species. As they say, nature works with what it's got, we've seen this with Foxp2, which shows up in derived form in humans, birds and whales (highly vocal species). Anyway, the abstract:
Here is an easier to digest piece in The Washington Post, Scientist s Find A DNA Change That Accounts For White Skin (the title isn't true of course, there are East Asians with white skin). And Nick Wade is on it of course. Thanks to Abhi & Theresa for the tip. You can find a PDF of the paper as "whiteskin" in the forums. Update: NPR has a nice summary. People seem interested in this topic. Addendum: Check out William Saletan's idiotic comment:
Saletan might have been tongue-in-cheek, but if he is he's only perpetuating public stupidity (something that doesn't need any reinforcement, thank you very much). First, less than half of the color variation even between Europeans and Africans is accounted for this gene, i.e., it is a necessary condition for "whiteness" (in the European sense), but it is not a sufficient condition. Living in an east coast metro area his model of a black-white dichotomy (i.e., the stories clearly suggest that East Asians lack the derived variant, but they aren't black to my knowledge) in any conception of race pops up (if ($race != white) { $race = black}). Second, it makes the standard conflation of skin color with populational identity. This map makes it clear that skin color tends to track latitude far more than the combination of latitude & longitude that would imply a tight correlation with geographical populations. If you step outside the black-white world skin color can not predict populational identity very well (except perhaps at the very fair end of the range where Northern Europeans are alone). And as for reaction #2, what idiots actually said anything like this? That is a contention so bizarre that it seems to have been produced by a 3rd grader (I'm expecting to falsified as to the expectation of chronological age). I wouldn't expect much from Saletan, but he writes the "human nature" column for Slate. How can I get a cushy gig like that??? Addendum II: Check out this post from a few years back on altitude adaptation to see what I mean about stumbling upon different fitness peaks via different strategies. Also, consider that many genes have pleiotropic effects, that is, they are implicated in a multitude of genetic pathways and influence many traits. There are many background assumptions operating when one says that locus x has fitness effect y on population z. Consider that perhaps light skin in the generality is a phenotype that is advantageous at higher latitudes, while dark skin is advantageous at lower latitudes. Since to the first approximation humans are creatures of Africa it makes sense that we would have fixed or constrained toward expression of genes that influence skin color toward a dark optimum. As non-Africans are a subset of Africans it seems plausible that the ancestral dark inducing forms of the genes will be shared. On the other hand, as the constraint for dark skin is released, because selection no longer favors it, the genes will start stumbling randomly in various directions via mutations. In the case of Europeans the MC1R locus seems to have walked in a random fashion and diversified greatly1 (30 alleles of greater than 1% frequency). On the hand, in East Asians the MC1R allele seems to have been selected toward one particular form that differs from the ancestral variant, in other words, constraint that limited the fitness toward those bearing the ancestral dark skin inducing allele(s) was released, but unlike Europeans selection now operated on a different allele and constrained diversification. This new finding makes that more intelligible: Europeans were given license to explore the range of MC1R variants because another locus was sufficient in hastening the induction of a light skin phenotype. Selection operates on genotype via phenotype, so the phenotype is sufficient to allow the individual to be fit and reproduce the genes, it is irrelevant what that particular conformation of genes that results in said phenotype is. An important point I am leaving out of this is that it is likely that MC1R has other fitness effects (recall the finding that redheads might be more sensitive to pain), and it is likely that this new locus is also implicated in other phenotypes, at least indirectly. In other words, loci don't explore the fitness landscape by their lonesome, but only in the context of changes and conditions on other loci. 1 - Negative frequency dependent sexual selection is another way to generate diversity. Hard to falsify though, and usually neutrality is assumed to be a legitimate null hypothesis. Labels: Pigmentation
Friday, August 19, 2005
It's really frustrating when you can't find information via google, but, it just reminds you how shallow the the data mining of search engine crawlers can be. On this weblog people have mentioned blondeness among Australian Aboriginals multiple times, and ultimately we really haven't gotten anywhere (no one has brought up novel data) because no one has any information to offer aside from what they read in C.S. Coon's books when they were younger. There isn't much out there on the web.
Luckily, I decided to check the local college library, and I found Joseph Birdsell's Microevolutionary Patterns in Aboriginal Australia, which has a large section addressing the issue of blondeness among the indigenous people of the antipodal continent. Below, I will summarize most of Birdsell's data and analysis so that google will at least have this to crawl now. But first, I want to address a minor point that often comes up. One hypothesis about Australian Aboriginal blondeness is that it is due to admixture with Europeans, in particular Dutch sailors who entered into undocumented liasons with native women prior to British colonization. This to me seems like a ludicrous assertion for the following reason: if the blonde alleles introgressed from another population, they can be thought of as proxies for the ancestral admixture of Western Europeans into these tribes. Though a very high frequency of tribal members exhibit preadult blondeness, there are almost no other European diagnostic phenotypes in evidence! That is, their skins are rather dark and their features classically Australian Aboriginal. Most people talk about European blondeness as if it is a recessive trait. I have issues with that simple idea, but, taking it at face value the frequency of blonde alleles in a panmictic population should be higher than the frequency of the blonde phenotype,1 so we are talking about a rather high level of admixture if the blondeness is due to European ancestry. On the other hand, there are no other visible signs of this ancestry. One could hypothesize of course that the initially low frequency (attained via admixture) spread through the population because of positive directional selection on the trait. So in that case the alleles are of European origin, but the frequency of blondeness is not diagnostic of ancestry because it is not a neutral trait. But Birdsell's data points away from a European origin for blondeness, and many of the recollections of readers of GNXP are correct as to the character of this trait among Australian Aboriginals. To review, there are two primary melanin pigments, dark eumelanin and red-gold pheomelanin. The dosage of these two pigments results in the various hair colors we see in people. Redheads tend to have a great amount of pheomelanin, but almost no eumelanin. Ash blonde people are the reverse when it comes to pheomelanin, while golden blonde individuals tend to be somewhere in the middle. People with auburn hair have relatively high levels of both. But note that pheomelanin is more diffuse and less abundant, and it is no a surprise that black haired individuals may simply mask their "red" pigment. Many people with black hair (including yours truly) go through a "red blonde" phase during hair bleaching, as the dense eumelanin granules are stripped away by the bleaching agents first. It seems that the expression of the phenotype is dependent on many genes, though a few, like MC1R, have an outsized influence (perhaps through regulation of other loci). This is probably one reason that despite the typological division of Europeans into "blondes," "brunettes" and "redheads," there tends to be a continuous gradation of color. Not only do the combinations of eumelanin and pheomelanin dosage add "mixed" categories (strawberry blonde, auburn) to the triplet, the expression of these pigments is not an "on" or "off" matter as one would expect if one locus was at the heart of the process. I have made the repeated argument that the "recessive" character of blondism and the "dominant" character of brunette hair is partially an artifact of how we classify hair color. All the various non-blonde hair colors, from brown to black are slotted into the "dominant" category, when I would argue that even among black haired people there is a wide variance of pigment concentration of eumelanin that visual inspection might miss, for example between a light skinned Japanese individual and someone from southern India or Africa (basically, one can not get below a certain level of reflective, so all the extra melanin does not register any change in color). Now, to the Australian Aboriginals. 1) The perception (based I assume in color plates in older anthropology books) that the blonde Aboriginals were ash in their coloration is correct. The reason, according to Birdsell, is that they exhibit very little pheomelanin in their hair. Of course there is a lack of eumelanin in the hair samples as well. Unfortunately Birdsell did not assay the concentration of granules quantitatively, but inspected them visually under a microscope. Nevertheless, he saw what was going on at the proximate level pretty well. It wasn't, to consider an outlandish example, a case where a yellow pigment was being produced that obscured the eumelanin. 2) There is both sexual dimorphic and paedomorphic tendencies to the trait. In short, pre-pubescent children are blonder, as are females. 3) This is not a rare trait that is expressed by a few individuals in many tribes. Rather, the frequency of the phenotype can approach 90-100% in children, and still remain significant even in adult males. Also, the "darkening" is often to a brown color, rather than black. 4) Birdsell suggests that the allele which causes this blondeness, in reality the loss of function or expression of both traits (dark and red pigment), is characterized by "incomplete dominance." The frequencies for the expression of the trait are extremely high. If it was a "recessive" trait the allele(s) must be close to fixed. I don't find his arguments persuasive because he didn't mention crosses between dark haired aboriginals and blonde aboriginals, in part because the unmixed peoples of this sort (that is, without European ancestry) are also not likely to go on cross-continental searches for husbands or brides from other Aboriginal groups. But, that being said, Birdsell offers the following observation: hybrids between Europeans and dark-haired (eastern) Aboriginals never exhibit hair that is lighter than brown. Obviously, not all Europeans are blonde, or carry blonde genes, but the conclusion of blonde phenotypic recessiveness is hammered home. Hybrids between blonde Aborigines and Europeans almost always exhibited the ash blonde phenotype of the Aborigines as children. I don't put too much stock in terms like "incomplete dominance," aside from that it is saying "hey, we don't know much about this gene." Nevertheless, I think the hybrid phenotype is a strong line of evidence that it isn't localized on the same part of the genome as the blonde loss-of-function alleles in Europeans. Crosses between dark haired Europeans and blonde Europeans do not almost always result in blonde children (many times the children are blonde and they become dark haired as they develop, but, Birdsell seems to suggest that inheritance pattern is more deterministic when one of the parents is an Australian Aboriginal blonde). 5) Birdsell notes that the blonde phenotype does not apply to all body hair. Almost all the rest of the body hair is rather dark, the only exception being the hairs on the forearm, which tend to be even blonder (that is, those who darken with adulthood retain blonde forearm hair). I would like to end with a tentative hypothesis. Obviously Birdsell is trying to convey the impression that this is a trait that is "incompletely dominant," even though it is a "loss of function" trait (eumelanin and pheomelanin seem to not be found in the hair). The "incompletely dominant" part suggests that there is a locus of large effect at work here. Additionally, Birdsell only mentions gradation in hair color as a function of development or maturation, not population. What I mean by this is that one doesn't get the impression of individuals with light brown or dark blonde shades as youth who become black haired as adults. Continuity (normalish distribution) is a feature of polygenic traits, while discrete or binary tendencies are exhibited by classical mendellian traits. With this in mind, I offer that perhaps these Australian Aboriginals carry an allele which results in the synthesis of a trans-acting factor which suppresses gene expression on the loci which control for melanin production (or, it could be interfering with a crucial regulatory step). This suppression is obviously dependent on factors relating to development and cell-cell differentiation, because the melanin is found in copious amounts in other body hairs as well as in the skin. A sequencing of the loci which we know affect melanin dosage might not turn up anything out of the ordinary in comparison to other dark skinned people. In contrast, I suspect many Europeans have multiple polymorphisms which result in the overall reduction in melanin production via melanocytes throughout their skin, their body hair as well as their irises. So why is this trait expressed in frequencies of 90%+ (that is, adults who started out ash blonde as youth) in the west-central deserts of Australia? Birdsell doesn't offer any selectionist reason, and I can't think of any environmental ones. There was obviously constraint on skin color, which makes sense in light of the protection that dark skin confers against radiation. The only thing I can come up with is sexual or social selection (ie; it might have been preferences for a particular type of child as opposed to males and females choosing each other for this trait). But it is basically a default hypothesis (I do not credit genetic drift in this case, but I do not know the demographic history of these tribes, so that is a possibility I suppose). Also, blondism might just be a byproduct of the allele's function, which we do not know yet (or, we know it, but have not made the connection). I was going to scan the map up, but I'm having some driver issues, so no go in that direction (if someone wants to find the book and scan it up and put it on flickr I will link to it-it's on page 196). Descriptively, you have a modal frequency of this phenotype in the middle of western Australia of 90-100%. The frequency drops off to around 50% by the southwest coast and the geographic center of the continent, and more sharply north toward Arnehm Land until the phenotype is almost nonexistent on the north coast. The phenotype is absent from the eastern third of the continent. Overall, one can imagine an area of the map where the phenotype is absent like a crescent, thick and rotund in the southeast, and becoming a relative sliver as it arcs around the zone of blondeness around its northern edge. Related: Black and strawberry. 1 - p2 + 2pq + q2 = 1. The "recessive" allele is usually signified by q. The q2 is the frequency of expression of the recessive phenotype, so for example, if the blonde allele is present in a frequency of 0.5 throughout the random mating population, 1/4 of the individuals will express it. If a population is 1/2 blonde, than 70% of the alleles floating in the population are blonde. So, if you had a tribe that was 50% blonde, if blonde alleles are neutral (no selective advantage), ignoring drift one could assume that 70% of the ancestry was European if the alleles had to have come from that source population. Of course, I don't think that the dominance-recessive concept really works well a lot of the time, and I certainly don't think that blondism is a one locus mendellian trait, contrary what they taught us in high school. Labels: human biodiversity, Pigmentation |
  Razib's Home Page GNXP Archives Interviews Blogroll Principles of Population Genetics Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism
Archives
July 2005 August 2005 September 2005 October 2005 November 2005 December 2005 January 2006 February 2006 March 2006 April 2006 May 2006 June 2006 July 2006 August 2006 September 2006 October 2006 November 2006 December 2006 January 2007 February 2007 March 2007 April 2007 May 2007 June 2007 July 2007 August 2007 September 2007 October 2007 November 2007 December 2007 January 2008 February 2008 March 2008 April 2008 May 2008 June 2008 July 2008 August 2008 September 2008 October 2008 November 2008 December 2008 January 2009 February 2009 March 2009 April 2009 May 2009 June 2009 July 2009 August 2009 September 2009 October 2009 November 2009 December 2009 January 2010 February 2010 Hello Movable Type archives August 11,2002 August 18,2002 August 25,2002 September 01,2002 September 15,2002 October 20,2002 December 08,2002 December 22,2002 December 29,2002 January 05,2003 January 12,2003 January 19,2003 January 26,2003 February 02,2003 February 09,2003 February 16,2003 February 23,2003 March 02,2003 March 09,2003 March 16,2003 March 23,2003 March 30,2003 April 06,2003 April 13,2003 April 20,2003 April 27,2003 May 04,2003 May 11,2003 May 18,2003 May 25,2003 June 01,2003 June 08,2003 June 15,2003 June 22,2003 June 29,2003 July 06,2003 July 13,2003 July 20,2003 July 27,2003 August 03,2003 August 10,2003 August 17,2003 August 24,2003 August 31,2003 September 07,2003 September 14,2003 September 21,2003 September 28,2003 October 05,2003 October 12,2003 October 19,2003 October 26,2003 November 02,2003 November 09,2003 November 16,2003 November 23,2003 November 30,2003 December 07,2003 December 14,2003 December 21,2003 December 28,2003 January 04,2004 January 11,2004 January 18,2004 January 25,2004 February 01,2004 February 08,2004 February 15,2004 February 22,2004 February 29,2004 March 07,2004 March 14,2004 March 21,2004 March 28,2004 April 04,2004 April 11,2004 April 18,2004 April 25,2004 May 02,2004 May 09,2004 May 16,2004 May 23,2004 May 30,2004 June 06,2004 June 13,2004 June 20,2004 June 27,2004 July 04,2004 July 11,2004 July 18,2004 July 25,2004 August 01,2004 August 08,2004 August 15,2004 August 22,2004 August 29,2004 September 05,2004 September 12,2004 September 19,2004 September 26,2004 October 03,2004 October 10,2004 October 17,2004 October 24,2004 October 31,2004 November 07,2004 November 14,2004 November 21,2004 November 28,2004 December 05,2004 December 12,2004 December 19,2004 December 26,2004 January 02,2005 January 09,2005 January 16,2005 January 23,2005 January 30,2005 February 06,2005 February 13,2005 February 20,2005 February 27,2005 March 06,2005 March 13,2005 March 20,2005 March 27,2005 April 03,2005 April 10,2005 April 17,2005 April 24,2005 May 01,2005 May 08,2005 May 15,2005 May 22,2005 May 29,2005 June 05,2005 June 12,2005 June 19,2005 June 26,2005 July 03,2005 July 17,2005 August 07,2005 Blogspot archives June 2002 July 2002 August 2002 September 2002 October 2002 November 2002 December 2002
10 questions for....
Parag Khanna James Flynn Jon Entine Gregory Clark György Buzsáki Heather Mac Donald Bruce Lahn A.W.F. Edwards Luigi Luca Cavalli-Sforza Joseph LeDoux Matthew Stewart Charles Murray James F. Crow Adam K. Webb Justin L. Barrett David Haig Judith Rich Harris Ken Miller Dan Sperber Warren Treadgold Armand M. Leroi John Derbyshire
Blogs
The GiveWell Blog Your Religion Is False Colby Cosh Steve Hsu Audacious Epigone Catallaxy Files Inductivist 2 Blowhards Genetic Future Agnostic Steve Sailer Dienekes Derek Lowe Razib Khan Razib at Comment is Free Secular Right Glenn Reynolds Jim Miller Kevin McGrew John Hawks Peter Fost Randall Parker Less Wrong Charles Murray Carl Zimmer EconLog Marginal Revolution
Principles of Population Genetics
Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism   Policies Terms of use © http://www.gnxp.com Razib's total feed: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
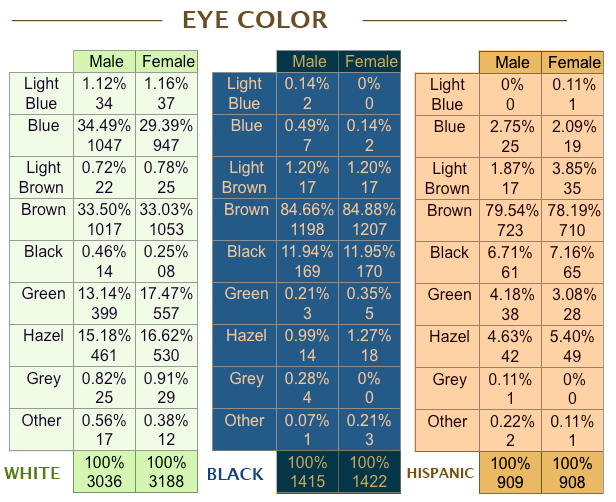
{kind=link}
{kind=link}
{kind=link}
{kind=link}