
|
Wednesday, December 16, 2009
How shellfish saved the human race:
Turns out, somewhere between 130,000 to 190,000 years ago, the human species was reduced to less than 1000 breeding individuals--just a few thousand people in total. Ancient, naturally driven climate change pushed our species to the brink, said Curtis Marean, Ph.D., a professor with the Institute of Human Origins and the School of Human Evolution and Social Change at Arizona State University. This is probably wrong. Though one of the many ideas about this period in human history have to be right. And there's always the tricky problem of falsification and testing alternative hypotheses when it comes to models of human evolution. From what I know humans are a relatively homogeneous species which underwent some sort of demographic expansion within the last 100,000 years. The tacit assumption seems to be that the proto-modern African lineage nearly went extinct, and then bounced back, possibly due to an exogenous shock. My own question: why couldn't it just be that a few bands simply exterminated or marginalized all the other African lineages? This is an assumption by many about modern humans as they expanded into Eurasia, more or less, so why not in Africa? Humans can certainly reproduce up to the Malthusian limit. In 1800 1.2 million humans were resident in New England, the vast majority of whom were descended from the 20,000-30,000 who arrived in the 1630s. And remember that the reproductive population is going to be a fraction of the census size. I could speculate on what gave a small subset of African humans an advantage, but I'll leave that to the comments. Labels: Human Evolution
Wednesday, November 11, 2009
Human-specific transcriptional regulation of CNS development genes by FOXP2:
...It has been proposed that the amino acid composition in the human variant of FOXP2 has undergone accelerated evolution, and this two-amino-acid change occurred around the time of language emergence in humans...However, this remains controversial, and whether the acquisition of these amino acids in human FOXP2 has any functional consequence in human neurons remains untested. Here we demonstrate that these two human-specific amino acids alter FOXP2 function by conferring differential transcriptional regulation in vitro. We extend these observations in vivo to human and chimpanzee brain, and use network analysis to identify novel relationships among the differentially expressed genes. These data provide experimental support for the functional relevance of changes in FOXP2 that occur on the human lineage, highlighting specific pathways with direct consequences for human brain development and disease in the central nervous system (CNS). Because FOXP2 has an important role in speech and language in humans, the identified targets may have a critical function in the development and evolution of language circuitry in humans. Ed Young has a long entry on this paper, along with context. Update: Someone on Twitter is suggesting we create a transgenic. Which direction? Labels: FOXP2, Human Evolution, Language
Tuesday, December 16, 2008
The last five chapters of R. A. Fisher's Genetical Theory of Natural Selection - about a third of the book - are devoted to human evolution. These chapters are seldom quoted and probably seldom read, even by Fisher enthusiasts. [Note 1]
There are some obvious reasons for this neglect. Much of this part of Fisher's book is concerned in a broad sense with eugenics, the very mention of which is sufficient to paralyse rational thought in some quarters. But even for those who are not scared of the e-word, there would be reasons for disregarding these chapters. The evidence on which Fisher relies is thin and out-of-date. His evidence on the heritability of human fertility, which is central to his arguments, depends entirely on studies of the British aristocracy, which is hardly a representative sample of the species. Apart from this, like many of his contemporaries (in the 1930s) Fisher believed that current fertility trends were dysgenic: that Britain (and other western nations) were threatened by a decline in the genetic quality of the population. For example, the psychologist R. B. Cattell estimated in 1937 that average IQ in Britain was falling at a rate of about 1 percent per decade. The snag with the dysgenic hypothesis is that the period since the 1930s has seen a large improvement in almost all measurable aspects of human 'quality': IQ, educational achievement, height, general health, and longevity. The average man or woman in Britain today lives about 20 years longer and has an IQ about 20 points higher than in Fisher's day (by 1930 norms). It is possible to argue, like Richard Lynn [Note 2], that an underlying genetic decline has been masked by an even larger environmental improvement, but from a practical point of view pessimists like Fisher and Cattell have been refuted by events. Nevertheless, there is much in Fisher's neglected chapters which is interesting and worth reading, and this post is intended as a brief taster.... Why Human Evolution is Special Fisher complains that the treatment of man in general works on evolution is usually superficial, and emphasises that human evolution is interesting and unusual enough to deserve extended treatment. He points out (p.192) that any animal that has undergone profound changes in its recent evolutionary history should be of special interest to the evolutionist. He mentions some of the more obvious special features of man - big brain, exceptional social organisation, use of artificially constructed tools, symbolic communication - and concludes that 'to the non-human observer mankind would present a number of highly interesting evolutionary inquiries and would raise questions not easily to be answered only by the use of comparisons and analogies'(p.192) . But the distinctive perspective he gives to human evolution is that, in contrast to most other species, natural selection in man operates mainly through differences in fertility, rather than mortality, and that it now operates (at least in 'civilized' man) with exceptional intensity (p.218, 228). He goes on to explore the interaction of social class, fertility, and sexual selection, and concludes that the combination of conditions producing an acceleration of evolutionary changes is 'peculiar to man' (p.269). Far from thinking that in modern man evolution has come to a halt, as some modern evolutionists (e.g. S. J. Gould, Steve Jones) have claimed, Fisher therefore believes that it is exceptionally rapid, and capable of producing significant changes even during recorded historical times. The Evolution of Fertility Fisher makes some brief but important general remarks on the evolution of fertility, which are applicable to all species, and not just to man. Despite this, his remarks have been generally overlooked. [Note 3] He argues that fertility, like any other trait, is subject to natural selection, and that the most important factor in determining optimal fertility is the amount of parental expenditure required: 'In organisms in which that degree of parental expenditure, which yields the highest proportionate probability of survival, is large compared to the resources available, the optimal fertility will be relatively low' (p.204). In 'civilized' man, the most important determinants of fertility are psychological. There are factors of temperament which determine the propensity to marry, whether marriage is early or late, and the degree of enthusiasm for children (p.210-13). But there are also social and institutional factors such as prohibitions on infanticide (p.218-21). These factors will themselves be affected by psychological influences which will vary over time, (p.219), since parents who are reluctant to commit infanticide will have more children surviving, and the children will tend to inherit their parents' temperament. Fisher argues that this is responsible for the changing historical views on infanticide, and minimises the role of religious doctrine, which itself (he argues) is responsive to the general mood of the population. In a splendidly Fisherian phrase he remarks: 'It would, I believe, be a fundamental mistake to imagine that the moral attitude of any religious community is to any important extent deducible from the intellectual conceptions of their theology (however much preachers make it their business so to deduce it)' (p.222) . Man versus Social Insects In several places (p.199-204, 271-2) Fisher compares and contrasts human and insect societies. He stresses the major difference that in insect societies reproduction is specialised in a reproductive caste, often with a single queen. An insect society therefore 'more resembles a single animal body than a human society' (p.200) and 'selection must in this case act exclusively on the reproductive insects via the prosperity of the societies from which they arise' (p.201). In the light of modern sociobiology this emphasis on the reproductive system may seem blindingly obvious, but in Fisher's time it was not, and even in the 1950s writers like A. E. Emerson still tended to neglect it. Fisher also has a most interesting comment on the origins of insect societies, suggesting that 'as soon as the young adults of any incipient social form took either to performing the preparatory labour for reproduction, or to tending the young, before they themselves had commenced to reproduce, the balance of selective advantage would have been shifted towards favouring the fertility of the foundress of the colony, and towards favouring equally the development of the organs and instincts of workers rather than of queens among her earlier, and possibly less well nourished, offspring' (p.205). In human societies, in contrast, reproduction remains individualistic, and genetic competition within communities is always present. Fisher does not entirely dismiss the importance of inter-group selection: 'Among small independent competing tribes the elimination of tribes containing an undue proportion of the socially incompetent, and their replacement by branches of the more successful tribes, may serve materially to maintain the average standard of competence appropriate to that state of society' (p.201). But even in this state of society competition within the community is present, and becomes more important as the size of groups increases (p.201). He later points out that 'The selection of whole groups is, however, a much slower process than the selection of individuals, and in view of the length of generation in man the evolution of his higher mental faculties, and especially of the self-sacrificing element in his moral nature, would seem to require the action of group selection over an immense period' (p.264. Incidentally, this is the first use of the exact phrase 'group selection' I have noticed in the literature. Sewall Wright, around the same time, uses 'intergroup selection'.) Fisher concludes that the main force in the evolution of such qualities has been individual selection, but powerfully enhanced by the action of kinship groups and sexual selection, which in the case of man also involves decisions by kinship groups. I will discuss this further in another post. To be continued, probably after Christmas..... Note 1: I will give page references to the easily available Dover edition (1958). There are no relevant changes from the first edition. Among Fisher's admirers, W. D. Hamilton does in his very first published paper refer to the 'human' chapters of GTNS (see Narrow Roads of Gene Land, vol. 1, p.8), but elsewhere does not, even when (as in his essay 'Innate social aptitudes of man') they would be highly relevant. Note 2: Lynn has written a book, Dysgenics, and various articles on this theme. He attributes the increase in average IQ (the Flynn Effect) mainly to improved nutrition. The awkwardness of his position is that his argument for dysgenic effects requires the genetic influence on individual IQ to be large, while his interpretation of the increase in IQ also requires the influence of environment to be large - larger in fact than the entire observed Flynn Effect, since this is the net result of a negative genetic trend and a positive environmental effect. This combination of requirements is not logically impossible, but it is uncomfortable. Note 3: the modern theory of the selection of optimal fertility is usually credited to David Lack, who gathered empirical evidence for it, but the key concept of optimal parental investment is contained not only in Fisher but in various other writers. Fisher himself credited the concept to Major Leonard Darwin. Labels: Burbridge, Human Evolution
Tuesday, December 02, 2008
The 10,000 Year Explosion: How Civilization Accelerated Human Evolution
posted by
Razib @ 12/02/2008 11:58:00 AM
 Just wanted to give everyone a heads up, Gregory Cochran's new book, The 10,000 Year Explosion: How Civilization Accelerated Human Evolution, is available for pre-order on Amazon. Of course, I wouldn't trust Amazon's publication date too much.... Just wanted to give everyone a heads up, Gregory Cochran's new book, The 10,000 Year Explosion: How Civilization Accelerated Human Evolution, is available for pre-order on Amazon. Of course, I wouldn't trust Amazon's publication date too much....Labels: Human Evolution
Wednesday, October 29, 2008
Gene Expression Profiles during In Vivo Human Rhinovirus Infection (also, ScienceDaily summary):
Rhinovirus infection significantly alters the expression of many genes associated with the immune response, including chemokines and antivirals. The data obtained provide insights into the host response to rhinovirus infection and identify potential novel targets for further evaluation.. About those viruses: Epidemiologists have established minimal population size and density thresholds for particular diseases (such as measels, mumps, rubella, smallpox, influenza, rhinovirus) to survive and spread. In small hunter-gatherer groups or even small farming villages, such diseases would have been incapable of spreading very far and woul have disappeared (Black 1975). This implies that many diseases must be recent. There's a reason that some of the cites for the adaptive acceleration theory use microbial models; here are lot's of them and they breed fast. Microbiologists are fond of reminding people that on the order of 90% of the cells in your body are bacteria resident in your gut; but I wonder if the last 10,000 years might not have been a boon for a whole host of virulent less friendly microbes which "tag along" with H. sapiens. Related: Toxoplasma gondii & human culture, Obesity germs, thrifty genes, Another Nobel for the New Germ Theory of disease and Toxoplasma gondii's South American origins and its influence on culture. Labels: Human Evolution, pathogens
Thursday, October 09, 2008
Yann reviews a recent paper, Lactase persistence-related genetic variant: population substructure and health outcomes.
Labels: Genetics, Human Evolution
Monday, October 06, 2008
Leading geneticist Steve Jones says human evolution is over. Steve Jones has an appointment in the Galton laboratory, and has written several books on human genetics (e.g., Y: The Descent of Man and Darwin's Ghost: The Origin of Species Updated). But he says things like this:
"Small populations which are isolated can evolve at random as genes are accidentally lost. World-wide, all populations are becoming connected and the opportunity for random change is dwindling. History is made in bed, but nowadays the beds are getting closer together. We are mixing into a glo-bal mass, and the future is brown." First, we're nowhere close to panmixia.* Second, there is going to be a large variance around the expectation. Even if you remove new mutations, there are a lot of variants out there for selection to pick up from the extant genetic background I would think. The future will not be brown for the same reason that people in an English village do not all have the same hair color despite there being a lot of intermarriage. Lots of other things to point to that leave you confused in that piece, but I'll leave it as an exercise for the readers.... * And what about the lack of importance of population size as a parameter effecting substitutions in Neutral Theory? I know there are ways you can object to this, but Jones' quote seems to garble many issues here. Update: I emailed an academic who I suspected would know if Jones was being quoted out of context or misrepresented. But they say that this is probably an accurate representation of his views (and they also seem to think that his coherency leaves a bit to be desired). Update II: Here is Chris Stringer's rebuttal to Jones: But Professor Chris Stringer, research leader in human origins at the Natural History Museum, London, said the idea that evolutionary pressures were no longer taking their toll on humanity was true of only western civilisation. *roll eyes* Someone should tell these guys that you don't need to die to not reproduce. Labels: Genetics, Human Evolution
Thursday, September 04, 2008
Multiple Advantageous Amino Acid Variants in the NAT2 Gene in Human Populations:
Data on the structure of haplotypes and their frequencies are compatible with a model in which slow-causing variants were present in widely dispersed populations before major shifts to pastoralism and/or agriculture. In this model, slow-causing mutations gained a selective advantage in populations shifting from hunting-gathering to pastoralism/agriculture. We suggest the diminished dietary availability of folates resulting from the nutritional shift, as the possible cause of the fitness increase associated to haplotypes carrying mutations that reduce enzymatic activity. Shorter version: a bunch of loss of function mutations which effect enzymatic activity in the ancestral genetic background shot up in frequency across disparate populations with the Neolithic revolution due to dietary changes. The paper is kind of a hodge-podge (e.g., not statistically significant, so let's hand wave :-). Also the appeal to balancing selection generally makes me suspicious...exactly how much balancing selection was their in our species? If there's a lot of it I assume it has more to do with frequency dependence and environmental heterogeneity than heterozygote advantage. Nevertheless, perhaps this is going to go down with amylase and lactase persistence as a recent adaptation driven by the shock of agriculture. I'm certainly not averse to the idea that nutritional deficiences due to the switch to agriculture drove a lot of recent human evolution. H/T Dienekes Labels: Genetics, Human Evolution
Significance Of Milk In Development Of Culture To Be Studied:
"The oldest pottery shards shown to contain milk were found in southeastern Europe, more precisely in what today is northeastern Greece. We believe that the mutation once grew common there and then became fundamental to the development of agrarian culture," says Anders Gotherstam, who will be coordinating the project. OK, if I'm reading this right...the hypothesis is that lactase persistence due to a mutation around LCT will be highest frequency in regions which experience agriculture, but haven't experienced it that long? In other words, perhaps the "end state" of agriculture societies is more like China, as milk culture gets squeezed on the Malthusian margins? If this is what they're saying I doubt I believe it. Labels: culture, Genetics, Human Evolution
Monday, June 02, 2008
Selection speculation: CLOCK and reward-dependence in Africans
posted by
agnostic @ 6/02/2008 02:39:00 AM
Since so many comments lead off with some variant of "I would guess," why not try to corrall them all into one post where they could serve a purpose? Each week I'll find some area of the human genome that shows signs of recent selection, see what phenotypes the gene affects, and although I'll likely provide the most convincing story, readers can conjecture to their heart's content about what might have driven selection. It may, for once, improve the discussion to comment while still deranged from last night's drink.
Let's start with the data: using Haplotter, we see that for the gene CLOCK, there is a signal of recent selection in Africans but not in Europeans or Asians. The CLOCK gene is involved in maintaining our circadian rhythm, and I started this search looking for between-group differences in being "day people" vs. "night people." However, other genes related to circadian rhythm -- PER1, PER2, PER3, Tim, CSNK1E, Cry1, and Cry2 -- show no signals of recent selection anywhere. So perhaps there is something else that CLOCK does that these others do not. It turns out that CLOCK is also involved in the dopaminergic system, which regulates mood and behavior. None of the OMIM entries for the other circadian rhythm genes mentions dopamine, mood, the reward system, etc. So it is more likely that CLOCK has undergone selection for its effects on mood and on the brain's reward system, rather than on circadian rhythm per se. In mice, and so potentially in humans as well, mutations in CLOCK make individuals lower in anxiety and higher in risk-taking (Roybal et al. 2007; free full text). Turning to data from Lynn & Martin (1995), which I turned into a convenient graph here, we see that Nigerians -- the African group that HapMap data come from -- are indeed the lowest in the world in the personality trait Neuroticism (the Israeli data-point must be a mis-coding by Lynn, who is known for doing so). They are also the highest in the world in Extraversion. Neuroticism measures a tendency toward anxiety and related states, while Extraversion measures, among other things, pace of living (manics living a faster-paced life) and excitement-seeking. So Nigerians show a psychological and behavioral phenotype that's fairly comparable to the mice with mutations in CLOCK, strengthening the hypothesis that selection at CLOCK has acted on personality rather than circadian rhythm per se. Now comes the fun part -- telling a story about why this phenotype was more advantageous in Western Africa than in Central Europe or Northeast Asia over the past 10,000 or so years. The low-anxiety and high-excitement-seeking phenotype is suspiciously like that associated with derived alleles at the DRD4 locus, and the novelty-seeking 7R allele at that locus does show up at intermediate frequencies in Africans (Chen et al. 1999; see a map here). I concur with Harpending & Cochran (2002) (free full text), who argued in the DRD4 case that the phenotype is more advantageous in "cad societies" than in "dad societies." More concretely, I suggest, based on the teachings of the pickup artist community, that low anxiety aids in approaching more females in a shorter time -- you are less paralyzed during the approach, and you get over rejection more easily -- and that higher risk-taking motivates you to approach more females. Contingent upon having other traits that are appealing to females (a muscular body, artistic skill, dancing or singing ability, a knack for flirting, and so on), the phenotype here would have increased male success in courtship of (multiple) females, a stronger determinant of reproductive success in a "cad society" than in a "dad society." Lastly, here are two testable predictions: 1) lowland South American and highland New Guinean tribes should also show signs of selection at the CLOCK locus, since they are even more prototypically "cad societies," and they have the highest frequencies of the 7R allele at DRD4. And 2) African Bushmen should show no signs of selection at CLOCK since they are more "dad societies" and have lower frequencies of the 7R allele at DRD4. (ALFRED does not have data on CLOCK.) The fact that Northeast Asians do not show signs of selection at CLOCK already supports the hypothesis, since they are more "dad societies" and have a very low frequency of 7R at DRD4 (and even the 2R allele, which shows up somewhat there, has a much more muted effect than does 7R). The lack of selection at CLOCK in Central Europeans is neither here nor there: they are more "dad societies" than Western Africa but not so much as East Asia; Europeans also have intermediate frequencies of 7R and DRD4. So we could have an example of "Rushton's Rule," where East Asians show no selection for low-anxiety / high-novelty-seeking alleles at either DRD4 or CLOCK, Europeans show selection at just one locus, and West Africans show selection at both. To the best of my knowledge, as they say, this is the first argument for why Africans show a signal of selection at CLOCK. Consider it part of the graduate student with a PC trend in studying human evolution. You heard it here first. References Chen, C., M. Burton, E. Greenberger, & J. Dmitrieva (1999). Population migration and the variation of Dopamine D4 receptor (DRD4) allele frequencies around the globe. Evolution and Human Behavior, 20(5): 309-324. Harpending, H. & G. Cochran (2002). In our genes. PNAS, 99(1): 10-12. Lynn, R., & T. Martin (1995). National differences for thirty-seven nations in extraversion, neuroticism, psychoticism and economic, demographic and other correlates. Personality and Individual Differences, 19(3): 403-406. Roybal, K., D. Theobold, A. Graham, J.A. DiNieri, S.J. Russo, V. Krishnan, S. Chakravarty, J. Peevey, N. Oehrlein, S. Birnbaum, M.H. Vitaterna, P. Orsulak, J.S. Takahashi, E.J. Nestler, W.A. Carlezon, Jr., & C.A. McClung (2007). Mania-like behavior induced by disruption of CLOCK. PNAS, 104(15): 6406-11. Labels: anthropology, babes and hunks, Genetics, human biodiversity, Human Evolution, Personality, speculation
Sunday, May 18, 2008
Male preferences and debunking myths about the evolution of the female form
posted by
agnostic @ 5/18/2008 06:49:00 PM
Click for Uncensored In the comments section to a 2blowhards post on booty shakin', blogger Alias Clio puts forth an argument from incredulity regarding several hypotheses I proposed: 1) that male preferences for different parts of the female body have, over time, correlated with personality traits; 2) that natural selection has had a role in causing some men to prefer one body part over another; and 3) that the correlation could be caused by some simple mechanism. She also repeats an evolutionary just-so story about why human females developed large breasts -- that is has something to do with face-to-face sex -- and that too is worth taking a hard look at (the story, that is). As to 1), the available data do paint a somewhat clear picture that assmen, boobmen, and legmen are not the same on average for personality traits. What Clio doubts is that the correlations here and now can be projected back into the past or into other parts of the world. That's true enough, but it's true of anything psychological, and only for technical reasons: when we discover which gene variants in males are implicated in preferring T over A, we can dig up or unfreeze ancient humans, sequence their DNA, and see if the males were boobmen or assmen. That's how we found out that some Neanderthals were probably red-headed, despite the fossil evidence not telling us anything about their hair color. Though everyone knows it's a dubious move, the best we can do to see what preferences may have been like in sub-Saharan Africa 100,000 years ago is to investigate present-day hunter-gatherers in Africa. The Hadza are a well studied hunter-gatherer group who live in Tanzania, and a study by Marlowe et al. (2005) (free PDF) shows that Hadza males prefer females who have a low waist-to-hip ratio in profile (i.e. due to protruding buttocks), rather than from the front (i.e. an hourglass or wasp-waist shape), while Western males prefer the converse. The authors did not collect personality data on the Hadza males, and did not test to see whether a male preferred boobs or buttocks, but in principle this would not be difficult to do, and we could see whether a similar pattern showed up among African hunter-gatherers. That brings us to 2), whether or not natural selection had a role in the emergence of boobmen. Clearly they are a new morph within homo sapiens. They are too high in frequency to be the result of de novo mutations here and now, and they did not all migrate from some pre-historic Martian colony of homo sapiens. That leaves genetic drift or natural selection. Genetic drift can cause allele frequencies to go up or down over time, but it cannot produce design. Mate preferences are too specific and coordinated during development to admit a believable drift explanation: natural selection appears to have fashioned them. But toward what end? We don't need to know, really. With the completion of the HapMap project, we are learning of tons of cases of natural selection in human beings, and we largely have no clue what it was up to. The numbers don't lie. Still, let's indulge in a little conjecture just to show that the idea isn't so perplexing in the case of boobmen. In many areas of life, there is no one best solution, and we face a trade-off. If I develop conspicuous ornaments, that may make me more attractive to females, but it may also give me away to predators more easily, or provoke the envy of duller looking males, who might ostracize me (no small matter in a social species). Duller looking males might avoid predators and envy-based ostracism, and may be able to work better in groups because of this, but they won't be as attractive physically to females. The result is that some fraction of males will be dull and the rest conspicuous. We would need tools from game theory and differential equation modeling to spell out what parameters are involved, and what the exact frequency of each would be at equilibrium. But the point is that neither is universally favored, so both will co-exist. So it could be with boobmen and assmen. I don't think these preferences per se were the target of selection, but again that they correlate with other personality traits that have been under selection. For instance, everyone says that compared to boobmen, assmen are more likely to have polygynous tendencies, to prefer short-term relationships, and to emphasize female qualities most relevant to the short-term (such as her most sexual body part, the derriere). We don't know if that's true, but it would be surprising if everyone had the same specific delusion. Since both short-term and long-term strategies have pros and cons, both could co-exist. If being a boobman is linked to a more monogamous orientation, we are asking how natural selection could have driven up the frequency of monogamous males in societies where boobmen are common, such as Northern Europe. Maybe agriculture there requires the father to stick around and provide for his kids, whereas in parts of sub-Saharan Africa where farming has lower energy requirements, females can farm on their own and not worry about whether the father will stay with her. I don't claim that this is the only way it could have happened; this example is just to illustrate how simple the process can be. Turning to 3), the mechanism does not have to be known in order to talk about the adaptive value of the trait (see Niko Tinbergen's Four Why's for clarification). We know that lighter eyes were selected for in Europeans, but we could know this fact even if we didn't know what biochemical pathways are involved in eye color. Still, let's indulge in a little more conjecture just to show how non-mysterious the mechanism can be. It may be as simple as testosterone level, with assmen having higher T than boobmen. This is an incredibly easy hypothesis to study empirically, though from Googling it looks like no one has done so. To repeat a finding from the boobman, assman, and legman study, though, the assmen and those who prefer both large breasts and large buttocks have more ambitious personalities and are Type-A businessmen. We also know that in various species, such as the dark-eyed Junco, higher testosterone makes a male more polygynous and less likely to stick around to help raise the kids. Whatever the mechanism turns out to be, investigating the matter is not so perplexing that we don't even know where to start looking. Last, let's examine some very popular but utterly ridiculous hypotheses for why human females evolved large breasts, summarized here. First of all, it is not true that human females have large breasts -- some have small, some medium, and some large. Look at the picture of the chimp in that summary -- you see human females with breasts that small (or large) all the time. This is not hairsplitting: it suggests that breast size reflects some trade-off. For example, the trade-off could be in fat deposition: if you have a fixed amount of fat and want to be conspicuous, you had better put the bulk of it in one place or the other. Only gifted (or cursed) females have so much to go around that they can have large breasts and large asses. Those who put it in their chests are probably pursuing a long-term mating strategy, and those who put it in their behinds are probably pursuing a short-term strategy, on the assumption that female supply has evolved to meet male demand. The evolution of breasts has nothing to do with mimicry of the buttocks -- can you think of any other way that a man might view buttocks-resembling things on a woman if he wanted to? Moreover, do assmen respond at all similarly to boobs as to the buttocks? This hypothesis predicts that they should be roughly interchangeable, but I don't even notice who has big or small boobs unless someone points it out to me, and I have no way of judging what "good boobs" look like, according to boobmen. It also has nothing to do with our species' face-to-face sexual position -- again, can you think of any other way a man might look at buttocks-resembling things while having sex? And as misleading as the name may be, doggy-style is not a trait that humans have lost, like a coat of body fur. Neither does it have to do with our bipedal posture: it's true that this posture would have obscured any rump swellings (as chimps have), but the fleshy buttocks have still been in plain view ever since -- and typically, more viewable from afar than the breasts, as they take up more volume. Five-hundred years from now, the scientific consensus will be that invoking bipedal posture as a driver of some clearly unrelated change was the 20th century equivalent of ancient Greek theories about trepidation of the spheres. Since such hypotheses are so easily debunked, why have they persisted for as long as they have? Napoleon said that you should never attribute to malice that which can be explained by incompetence, and here the incompetence surely stems from the majority of researchers and commentators in this area being boobmen, legmen, or women. (Women will grudgingly admit that boobs and legs can be things of beauty, but recoil in disgust upon pondering the booty.) We all have a tendency to extrapolate from the personal to the universal, but when I find out that someone thinks or behaves differently than I do, I ask what forces could cause them to think or behave in such deviant ways. That's how you get a clearer picture of how the world works, but it relies on there being a diversity of views within the field. It's about time some assmen joined the ranks of sex researchers to set the field straight. Labels: babes and hunks, Human Evolution
Sunday, February 17, 2008
Human Culture Subject To Natural Selection, Study Shows:
The Stanford team studied reports of canoe designs from 11 Oceanic island cultures. They evaluated 96 functional features (such as how the hull was constructed or the way outriggers were attached) that could contribute to the seaworthiness of the canoes and thus have a bearing on fishing success or survival during migration or warfare. The study is coming out on the 19th in PNAS (so that means it will show up on the website at some time after that date). As most of you know in the 1960s the neutral theory of molecular evolution emerged in response to the finding that there was a great deal of extant genetic variation on allozyme loci (OK, to be fair neutralist ideas predate the empirical results; but I think it is clear that those results made the model intellectually far more compelling). Prior to this there were two broad schools of evolutionary genetic thought; one group accepted that there would be low levels of polymorphism due to balancing selection, and another assumed that there would be little to no polymorphism because of selective constraint. No matter the rearguard attempts by the likes of Richard Dawkins to argue that molecular variation "doesn't count," I think the neutralist (or nearly neutralist) insights are important in giving us a better understanding of the nature of evolutionary dynamics on the genomic scale. In The Origins of Genome Architecture Mike Lynch argues that low effective population sizes have had a strong role in shaping the character of genomic variation in more complex organisms. In other words, we are all non-adaptationists now! What does any of that have to do with the paper above? Peter Richerson & Robert Boyd, L. L. Cavalli-Sforza and Marcus Feldman and E. O. Wilson & Charles Lumsden have all attempted to show how evolutionary processes are relevant to our understanding of human soceties. Unfortunately, as L. L. Cavalli-Sforza observes, cultural anthropologists are less interested in understanding humans as opposed to interpreting them. Formal frameworks to accompany the mass of empirical observations are simply neglected or seen as unnecessary. This is an unfortunate overreaction to the hubris of earlier generations of anthropologists who attempted to shoehorn all human variety into a set of functional adaptations. Instead of a happy medium where skepticism is balanced with empiricism and rationalism, anthropology has swung from a total lack of critical analysis toward one where positive assertions are eschewed on principle (unless, of course, those assertions are directed toward Western culture). In Darwin's Cathedral David Sloan Wilson tries to make an argument for resurrecting a functional understanding of cultural traits as adaptations. I think that this sort of work is hard-going, at least beyond the level of triviality (e.g., the rationales for why the Inuit dress the way they do is rather straightforward). That is because "culture" is a very broad and ill-defined term and the selective pressures are myriad; the environment, the social matrix and the correlations with other traits are all critical. Wilson's methodology in Darwin's Cathedral was to use case studies; I don't think that that will cut it. Rather, massive surveys of collected data tested via statistical methods are probably more useful in extracting out the adaptive trends as a function of time and space. I do not, for example, think it is a coincidence that over the last 2,500 years all the complex cultural traditions on the World Island became associated with what we would call "Higher Religions," roughly, the fusion of supernaturalism with philosophy and institutional structures. But were these parallel developments a function of the specific adaptive needs of these complex societies? Or where they perhaps inevitable byproducts of the sufficient intersections of modal human psychology with the rise of the novelties of mass post-tribal society? These are big complex questions. I think that are certainly functionally significant cultural adaptations. That being said, I am not sure sure that they are responsible for the preponderance of between cultural variation. To go back to the example of Higher Religions, I think one can plausibly argue that some sort of synthesis between intuitively appealing extant supernaturalism with the intellectual & institutional abstracting tendencies of complex societies made them inevitable, necessary perhaps. Societies which were united by a common religious ethos may very well have been more fit than societies still characterized by a welter of tribal gods uncomfortably corralled under one political dispensation (though the dynamic might usually have been played out within an intrasocietal context; e.g., the introduction of Buddhism to Tibet and Japan by a particular faction at court and the subsequent nativist reaction with failed). But the specific nature of the Higher Religions may very well be arbitrary, neutral so to speak, because like a synonymous substitution they have no functional significance. Obviously the paper above targets the law hanging fruit. Engineering is not contingent upon the caprice of human social dynamics; it works, or it doesn't, by the grace of Mother Nature. But it's a start, as it is a reality check upon those who would argue that the full sample space of cultural possibilities are theoretically at play, and equally likely. The next step is to start examining traits not so strongly constrained by physical conditions. Labels: culture, Human Evolution
Monday, January 14, 2008
Back in the days before I'd ever read any probability or population genetics, I imagine I considered, as many laymen still do, evolution as a sort of deterministic march towards some optimum. I still remember being amazed at the simple equations that show how much stochasticity is involved; how random chance and historical accident can shape the fate of genetic variants. But are there cases where the layman's instinct is correct, where we can say that evolution was deterministic? Obviously, in some sense this is impossible to prove; one can't simply rewind the clock a thousand times and watch the outcomes. But there are natural experiments that I think shed some light on the subject.
The advent of dairy cultures in various human populations around the world provides one such natural experiment. I'm writing about this because of a recent study identifying yet another allele leading to lactose tolerance, this time in a Saudi Arabian population that drinks sheep's milk. A previous study, regular readers may remember, identified three other polymorphisms leading to the phenotype in Sub-Saharan pastoralists. Along with the "European" allele, this brings the total of probable lactose-tolerance-causing mutations segregating in humans to five. Let's make some assumptions: lactose tolerance is perfectly dominant, has a selection coefficient of around 0.1, and all these mutations will continue to fixation (this last one would be almost certainly true if the selection coefficient were constant--all the alleles have escaped the stochastic phases of their trajectories--but is an open question. What is the fitness advantage today of lactose tolerance? Surely this is testable). With these assumptions, one predicts that lactose tolerance has arisen around 25 times since it became advantageous. Given that we're talking about less than ten thousand years since dairy farming, that's quite remarkable. The relevant parameter here is the mutational target size--if lactose tolerance could only be caused by a change at one particular base pair in humans, it would never have arisen independently so many times. But with a mutational target so large, and a selection coefficient so strong, it becomes inevitable that any culture that developed dairy farming would eventually develop lactose tolerance. But it still seems amazing to me that it happened so quickly! Labels: Genetics, Human Evolution, Population genetics
Tuesday, January 01, 2008
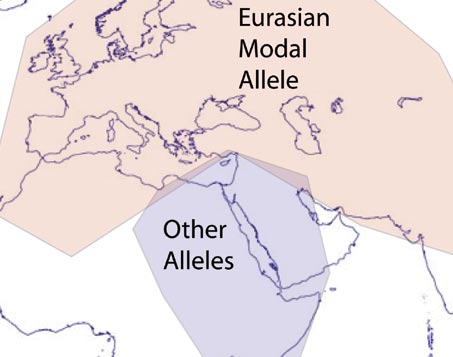 Evidence of still-ongoing convergence evolution of the lactase persistence T-13910 alleles in humans makes the case for a common Eurasian group of lactase persistence alleles across Eurasia, and among some African populations, derived from one mutational event within the last 10,000 years. A novel polymorphism associated with lactose tolerance in Africa: multiple causes for lactase persistence? points to other possible evolutionary events. The use of the term Africa is somewhat deceptive, these other alleles are found in groups like the Bedouin Arabs. One assumes that with the long history of agriculture in the Middle East these local alleles would have been relatively ancient as well. Why didn't they spread north as agriculture spread? And why didn't they spread east with agriculture? More of a question at this point. Evidence of still-ongoing convergence evolution of the lactase persistence T-13910 alleles in humans makes the case for a common Eurasian group of lactase persistence alleles across Eurasia, and among some African populations, derived from one mutational event within the last 10,000 years. A novel polymorphism associated with lactose tolerance in Africa: multiple causes for lactase persistence? points to other possible evolutionary events. The use of the term Africa is somewhat deceptive, these other alleles are found in groups like the Bedouin Arabs. One assumes that with the long history of agriculture in the Middle East these local alleles would have been relatively ancient as well. Why didn't they spread north as agriculture spread? And why didn't they spread east with agriculture? More of a question at this point.Labels: Genetics, Human Evolution
Monday, September 17, 2007
John Hawks has a series of great quotes from a 1963 article by Dobzhansky on the interplay between genetic and cultural evolution. How about this:
Being an anthropologist only by avocation, I may perhaps venture to claim for anthropology more than most anthropologists dare claim for themselves. The ultimate function of anthropology is no less than to provide the knowledge requisite for the guidance of human evolutionRather ambitious, no? Labels: Genetics, Human Evolution
Wednesday, August 08, 2007
I'm as much of a sucker for the flawed thought experiment as the next guy, so I'll pass on this one, via Robin Hanson:
Scott Aaronson asks a great question:Hanson's answer is far too simple a selective scenario. I'm even inclined to think that the evolution of sexual jealousy and the evolution of "justice" are less different than the question presupposes. It's easy to say "imagine a situation where stealing from an individual would lead to their starvation", but less easy to actually imagine it. How different were food and sexual access to our ancestors? The answer, I'm inclined to think, is: not so different. Among chimpanzees, food and sex are both commodities to be traded: Political coalitions were recognized early on as part of an elaborate 'marketplace of services' in which chimpanzees trade grooming, sex, food and support. The rules of reciprocity governing social exchange are only beginning to be understood, but evidence is accumulating that chimpanzees repay both positive acts (for example, sharing food preferentially with previous grooming partners) and negative acts (for example, squaring accounts with those who previously opposed them) [citation]While it's difficult to study these sorts of exchanges, a specific instance of male chimpanzees exchanging sexual access to females for political support was recently documented [citation], suggesting that perhaps this moral instinct Hanson is ready to justify evolutionarily isn't quite so universal. That's all in chimpanzees, of course, and it's true we live in a society where rape is certainly considered more reprehensible than stealing (of course, rape in wartime was considered par for the course until very recently). I'd be interested to see a study, however, comparing punishments for rape versus stealing across different societies, ranging from hunter/gatherer to our own. Is stricter condemnation of the forceful taking of sex as compared to the forceful taking of food a human universal? Labels: Evolutionary Psychology, Human Evolution
Friday, August 03, 2007
Note: I got this article via AJHG's RSS. It doesn't seem to have gone live on the site so there might be temporary problems accessing the link.
Evidence of Still-Ongoing Convergence Evolution of the Lactase Persistence T-13910 Alleles in Humans: A single-nucleotide variant, C/T-13910, located 14 kb upstream of the lactase gene (LCT), has been shown to be completely correlated with lactase persistence (LP) in northern Europeans. Here, we analyzed the background of the alleles carrying the critical variant in 1,611 DNA samples from 37 populations. Our data show that the T-13910 variant is found on two different, highly divergent haplotype backgrounds in the global populations. The first is the most common LP haplotype (LP H98) present in all populations analyzed, whereas the others (LP H8-H12), which originate from the same ancestral allelic haplotype, are found in geographically restricted populations living west of the Urals and north of the Caucasus. The global distribution pattern of LP T-13910 H98 supports the Caucasian origin of this allele. Age estimates based on different mathematical models show that the common LP T-13910 H98 allele (~5,000-12,000 years old) is relatively older than the other geographically restricted LP alleles (~1,400-3,000 years old). Our data about global allelic haplotypes of the lactose-tolerance variant imply that the T-13910 allele has been independently introduced more than once and that there is a still-ongoing process of convergent evolution of the LP alleles in humans. Two things to note 1) A recent common origin for much of the Eurasian lactase persistence phenotype is interesting. The period between 5 and 12 thousand years ago was obviously very significant and an inflection point in many ways in terms of the history of our species. Surveys of neutral markers which are supposedly reasonable proxies for ancestry imply that we should be cautious about mass population replacements across Eurasia. For example, it seems likely that the majority of Europeans and South Asians are descended from lineages already extant within their current geographic bounds at the end of the last Ice Age (though more or less significant impacted by population waves of advance triggered by the Neolithic revolution in the Middle East). Nevertheless, some biologists have argued that the sweeping action across demes of mutations of large effect are powerful enough to maintain species continuity and drive broad phenotypic convergences. One can conceptualize the genetic dynamics at work as a broad substrate of ancestrally informative alleles clustered across Eurasia, but tightly laced together by synchronous sweeps and pulses of functionally salient genes. 2) The newer, localized, LP variants are intriguing. The time window is very narrow here. Evolutionary theory tells us that in reaction to a strong selective force phenotypic change may immediately be affected by mutations of large effect. These mutants may be good at what they are meant to do, but also have negative side effects. Over time various selection pressures will reshape the genetic architecture with a host of modifiers and smaller effect mutants which result in a population subject to less stress via correlated responses due to the initial mutant of large effect. I wonder if the newer variants are signals that the genetic background is still working to optimize adaptation to a high lactose diet. Related: See here and here. Labels: Genetic, Human Evolution
Monday, June 25, 2007
Humans Have Spread Globally, and Evolved Locally:
No one yet knows to what extent natural selection for local conditions may have forced the populations on each continent down different evolutionary tracks. But those tracks could turn out to be somewhat parallel. At least some of the evolutionary changes now emerging have clearly been convergent, meaning that natural selection has made use of the different mutations available in each population to accomplish the same adaptation. Nothing new to readers of this weblog, but Wade does a good job surveying the various angles. Related articles on recent human evolution. Labels: Evolution, Genetic, Human Evolution
Sunday, June 24, 2007
Recently Razib posted on a review of sperm competition (PDF), part of which claims:
Kilgallon & Simmons [(2005)] documented that men produce a higher percentage of motile sperm in their ejaculates after viewing sexually explicit images of two men and one woman (sperm competition images) than after viewing sexually explicit images of three women. Motile sperm are the kind that are capable of moving themselves by swimming. The idea is that if a guy watches a video with two guys and one girl, he'll try to leave more swimming sperm since they now have to compete with those of the other man (so his mind thinks). Now, the proper reaction is to think that two guys going at one girl simultaneously was probably as rare, or even more rare, in our evolutionary past than it is today, so how would we show an adaptation to it? Fortunately the study in question is available online for free -- here -- and it's only two pages long, so read it. Let's review the key findings. In the Results, we read: Subjects viewing images of sperm competition had a greater proportion of motile sperm in their ejaculates than those viewing images of females (52.1 +/- 7.3% versus 49.3 +/- 8.0%; F1,23=5.08, p=0.034). That 3% difference in means is swamped by the standard-error bars of 7% - 8%, and the p-value, while under the 0.05 threshold, is uncomfortably close to it, considering the margin of error and the smallish sample size. So this result could easily be a fluke -- or not, but it warrants no confident statement that the study "documented" a pattern. "Somewhat suggested," perhaps, but gimme a break. This equivocal data aside, it gets worse when the authors looked at the concentration of sperm per volume: Men viewing images depicting sperm competition had fewer sperm in their ejaculate than those viewing images of females (61.35 +/- 1.27 versus 76.64 +/- 1.26 * 10^6 sperm ml^-1; F1,36=8.48, p=0.0061). [my emphasis] Now the effect is clear, but it contradicts the hypothesis that men's bodies will make more of an effort to defeat the other male's sperm when watching "sperm competition" images. In fact, given these two findings, maybe "two guys on one girl" ought to be called "sperm anti-competition" images. The strongest finding (read the paper for the data) was that men who thought the pornography was "more explicit" than what they'd seen previously had a much higher percentage of motile sperm and much higher concentration of sperm per volume. The authors suggest that men for whom this sort of pornography was old news had become habituated to it. That rings true anecdotally: I'm sure most guys recall how loudly their heart was pounding when they watched their first adult film. This raises an interesting possibility: if it's largely the novelty factor that's causing men to produce more sperm, and more motile sperm, would this carry over into the case where novelty was based on the ethnicity of the girl? The study design would be pretty simple: recruit a bunch of Latino-American and Anglo-American men, and randomly assign them to two groups, one that watches a scene featuring one man and one woman of the same ethnicity as the viewer, and another that watches a scene featuring one man of the same ethnicity as the viewer and one woman of the opposite ethnicity (Latin or Anglo). Then see if "jungle fever" played any role in how much sperm the men produce. I'm assuming the actresses in all scenes would be rated beforehand to ensure that any guy would find them very attractive. That's why it has to be Latinos and Anglos -- Blacks and Anglos might not work simply because too many White men don't find Black women as attractive as women of other groups, and that could be a problem. The other reason you might expect guys to produce more sperm when viewing a different-ethnicity girl is that throughout most of human history, including today in most parts of the world, it was incredibly rare to see someone from a noticeably different ethnic group. Even in the few cases where it happened, one of them would have been "just passing through" the other's region. Thus, the male would not know anything about her mating habits, and would have to assume the worst -- that she had a mate already. And if such encounters were fleeting, "strangers in the night" situations, then he would stand to gain everything by impregnating her. He'd never see her again, after all, so why not go the extra mile sperm-wise to make sure? Then he'd have another man raising his child: all benefit and little cost. You could also look at sperm content from real-life interracial couples, but there could be confounding factors. Maybe if you conducted the research where there were hordes of youngsters who were more-or-less open to mating with anyone -- say, Cancun during Spring Break -- you might be able to collect enough subjects to randomly assign them to the real-life versions of the video study suggested above. "Y'know, as long as you're mating with any old person, why not take part in our study?" Labels: Human Evolution, man-goo games
Saturday, June 16, 2007
Dissecting the regulatory differences between human and chimp
posted by
p-ter @ 6/16/2007 04:08:00 PM
Many evolutionary biologists are content to focus on large-scale trends in evolution-- they debate whether protein-coding or regulatory changes are more important in adaptive evolution, or look to compare the numbers of genes undergoing selection in different lineages. To each their own, of course, but I find those sorts of questions unsatisfying (for the moment), because ultimately, they cannot be answered without a molecular, reductionist approach. That is, the major breakthroughs in evolutionary biology will be in identifying specific molecular changes that have occurred during evolution and characterizing their phenotypic effects.
Case in point: the study of gene expression differences between humans and chimps. There have been a number of studies on the general trends in gene expression evolution between the two species. Some tissues are more constrained, some tissues less so. This is of interest, of course, but what we (I?) really want to know is: which specific genes are differentially regulated in humans as compared to chimps, in which tissues, when in development, and what specific genetic changes cause this differentil expression? The large-scale evolutionary trends will become apparent once this (extremely difficult) question is answered. It's because of the apparent intractability of this question that people tend to shy away from it. However, that's starting to change. Last year, using a painstakingly constructed multi-species microarray (as opposed to the human specific one used-- to study both humans and chimps-- in previous studies), Yoav Gilad identified specific genes in the liver (chosen because of its relative homogeneity) that have undergone human specific regulatory changes. Now, he's looking for the specific polymorphisms underlying these changes. A recently published pilot study explains the approach: Most phenotypic differences between human and chimpanzee are likely to result from differences in gene regulation, rather than changes to protein coding regions. To date, however, only a handful of human-chimpanzee nucleotide differences leading to changes in gene regulation are known. In order to identify differences in regulatory elements between human and chimpanzee, we focused on ten genes that were previously found to be differentially expressed between the two species. We then designed reporter gene assays for the putative human and chimpanzee promoters of the ten genes. Of seven promoters that we found to be active in human liver cell lines, human and chimpanzee promoters had significantly different activity in four cases, three of which recapitulated the gene expression difference seen in the microarray experiment. For these three genes, we were therefore able to demonstrate that a change in cis influences expression differences between humans and chimpanzees. Moreover, using site directed mutagenesis on one construct, the promoter for the DDA3 gene, we were able to identify three nucleotides that together lead to a cis regulatory difference between the species. High-throughput application of this approach will provide a map of regulatory element differences between humans and our close evolutionary relatives.Something to look forward to... Labels: Genetics, Human Evolution
Monday, May 07, 2007
So, I found this this strange press release in my RSS feed. Here's the important bit:
Gene sequencing revealed a mutation specific to humans that triggers a change in the splicing pattern of the neuropsin gene, creating a new splicing site and a longer protein. Introducing this mutation into chimpanzee DNA resulted in the creation of type II neuropsin. "Hence, the human-specific mutation is not only necessary but also sufficient in creating the novel splice form," the authors state. The full article is titled Human-Specific Mutation Leads to the Origin of a Novel Splice Form of Neuropsin (KLK8), a Gene Involved in Learning and Memory, and it is in a journal called Human Mutation. There seems something really stilted in the English in the press release, and the claims feel way too grandiose, but I figure if there is a 1% chance this is accurate I'll throw up a post. Those of you with neuro chops will probably sniff out the crap earlier then I.... Labels: Human Evolution
Ancient Australians Were a People Apart. Money shot:
A team headed by Georgi Hudjashov of Tartu University in Estonia analyzed variation in the Y chromosome and mitochondrial DNA (mtDNA, which is inherited only from women) in several hundred people from Australia and New Guinea. By knowing the mutation rate of mtDNA and comparing Australian samples with those from Asian populations, the team ascertained that the Australian and New Guinean populations branched off from a parent population 50,000 years ago, and that no significant additions to the aboriginal Australian gene pool had been made until modern times. "Australia was colonized, then nobody else came," says co-author Peter Underhill of Stanford University in Palo Alto, California. The study will be published in PNAS this week. The article notes that there seems to have been morphological evolution in Australia over the past 50,000 years (e.g., from gracile to robust). Some have adduced from this that waves of migration of disparate peoples. But, remember, phylogeny does not necessarily track morphology over evolutionary time scales. In Dragon Bone Hill two paleoanthropologists make the argument that the robust physique of Australian Aboriginals was a response to selective pressures derived from intercommunal melees (!?!?! You read that right!). Whatever the truth, I don't doubt that 50,000 years is long enough a time period that in situ selection upon extant genetic variation could reshape the modal morphology of populations. Additionally, we know that Australia was an not island ark totally separate from the rest of humanity, dingos were introduced within the last 10,000 years. It seems almost certain that the northern coast of Australia was occasionally visited by sailors from the Indonesian archipelago (for supplies such as water). Let's take at face value the results derived from these two neutral lineages and assume it is an accurate representation of total genome content in regards to ancestry. Explicitly, the vast majority of lines of ancestry of Australian Aboriginals today coalesce back to a founding population which reached the continent's shores 50,000 years ago. That does not negate the possibility that alleles may have crossed from the Eurasian mainland and so driven adaptive evolution amongst Australian populations. Just as they picked up the dingo from traditions of canine domestication originating in Eurasia, so Aboriginals may exhibit a non-ancestral genetic signature on many loci which are phenotypically salient. Labels: Human Evolution
Wednesday, March 21, 2007
Over at her website Judith Rich Harris has posted her article, Parental Selection: A Third Selection Process in the Evolution of Human Hairlessness and Skin Color. When I asked Judy 10 questions I expressed some skepticism about this theory:
4) In your 2005 response to the Edge Question, "What Do You Believe Is True Even Though You Cannot Prove It?," you alluded to two things, 1) selection for light skin 2) hairlessness by parents in infants. When you pointed to these facts, did you do so in light of recent genetic work which suggests that dark skin might have evolved in humans as a response to loss of body hair? In other words, one trait would never been selected for if not for the other. Here is the abstract from the artice: It is proposed that human hairlessness, and the pale skin seen in modern Europeans and Asians, are not the results of Darwinian selection; these attributes provide no survival benefits. They are instead the results of sexual selection combined with a third, previously unrecognized, process: parental selection. The use of infanticide as a method of birth control in premodern societies gave parents - in particular, mothers - the power to exert an influence on the course of human evolution by deciding whether to keep or abandon a newborn infant. If such a decision was made before the infant was born, it could be overturned in the positive direction if the infant was particularly beautiful - that is, if the infant conformed to the standards of beauty prescribed by the mother's culture. It could be overturned in the negative direction if the infant failed to meet those standards. Thus, human hairlessness and pale skin could have resulted in part from cultural preferences expressed as decisions made by women immediately after childbirth. First, on a pedantic note, let me lodge my general protest in regards to the assumed decomposition of natural, sexual, and, Judy's putative third factor, parental selection. I'm reading some cognitive psychological work now about categorization and one thing that struck me as very apt in regards to how humans conceptualize the world is that we're always trying to break nature apart its joints, so to speak. Most regular readers know I'm a fan of consilience, and so I must reiterate and be a nag about the fact that many dynamic selective processes may bound in nature, but fundamentally they're all of a piece. As J.B.S. Haldane said: fitness is a bugger, naming selection is easy, characterizing it both accurately and precisely can often be very hard. The gene is the unit of selection, the various levels and dimension are all equal under God's eye. In any case, to the thrust of the hypothesis. First, in the past few years a lot of work has been done on skin color. Last year I observed that Armand Leroi's afterward to Mutants where he notes that we don't understand skin color will need a coda in future editions. Here's what we know, so far.... 1) In the 1960s human geneticists using classical pedigree analysis determined that 4-5 loci, genes, explained most of the intergroup variation between blacks and whites in regards to skin color. 2) In the past few years genomics has generally confirmed this view, there are a few loci of large effect (e.g., SLC24A5 in Europeans vs. non-Europeans) which explain intergroup differences in complexion. 3) But, equivalent phenotypic values can be attained via alternative genetic architectures, and this seems to have happened. In other words, light-skinned northeast Asians are not necessarily light for the same reason that Europeans are. Even if the change occurs upon the same locus, the allele or haplotype may be different. 4) Different evolutionary dynamics might affect the various loci. For example, in Europeans MC1R is highly polymorphic, suggesting either diversifying selection (e.g., frequency dependence) or a deep coalescence time (perhaps MC1R built up a great deal of variation during the residence of hominid lineages in Europe and modern humans simply assimilated the local depigmentation alleles?). 5) Also, some of these loci under selection seem to be relatively recent (e.g., perhaps within the last 10,000 years). Like LCT (lactase persistence) they leave a powerful imprint on the genome via a selective sweep. How does this square with the hypothesis presented above? First, some prelims. Selection of all sorts can be hard to get a grip on. After all, we bandy about selection coefficients, s, in a vacuum, when they vary within their environmental contexts. Environment in this case can mean the natural world, the social world and other genetic parameters (via genetic interactions). Negative frequency dependence also throws a monkey wrench into these processes by making the selection coefficient a function of the frequency of the traits. But we need to start from simple models and build up in complexity. The example of the !Kung woman who did not want to kill her light-skinned daughter is illustrative of Judy's hypothesis, but, it is simply a starting point. Nevertheless, I think it highlights a weakness: the genomic data is shedding light on the possibility that selection for loci which cause light skin (or, more properly explain a proportion of the intergroup variance) occurred long after the first humans settled the temperate zone. If the parental preference for light skin (which derives from the deep seated sensory bias which is also the root of sexual selection) existed prior to the arrival in the northern latitudes why is it that Eurasian populations seem to exhibit pulses of selection relatively late in history? One could make the argument, assuming that parental and sexual selection were paramount, that child and mate choice were simply not operative prior to this time period. Sexual selection works ideally through polygynous mating systems where there is a great deal of reproductive skew. Peter Frost has argued that blondeness in European females emerged through a form of sexual selection where males selected from a finite sample space of females because of the nature of the low latitude tundra, but the operative principle is the same, selection upon heritable variation. Perhaps within the last 10,000 years large boom and bust cycles in populations through the World Island has resulted in truncation selection events which reshaped the genomes (and generated high selection coefficients who show up in the long haplotypes)? I really don't know, but, I think one must say that it is more complex and contingent than a simple relaxation of functional constraint and the operation of innate preferences. Additionally, I don't see much exposition of the details of sexual selection theory in Judy's article. For example, runaway sexual selection occurs very fast, and is often quickly checked by functional constraints. I believe that her hypothesis about light skinned preference being very deep implies some sort of sensory bias. The intersexual difference in coloration is obviously real. There are biological reasons for this (hormone levels), but the cultural amplifiers of this tendency are rather universal. On this point the hypothesis is on strong ground, civilizations throughout the world seem to value female (relative) paleness. But, I think this point still goes back to my previous issue in regards to time scale: is this a phenomenon of the mass societies? In any case, I'll leave it at that. There's a lot to chew on here. My main point of contention with this hypothesis is that I think the time depth is off. The most current results out of genomics suggest repeated and independent evolutionary sweeps in northern Eurasia at various times generating the phenotype of light skin. If the preference for light skin is deep within our natures, even predating bipedalism, it seems that it should have manifested immediately with the move of H. sapiens sapiens to the northern latitudes 30,000 years ago. Labels: Evolution, Genetics, Human Evolution, Pigmentation
Monday, February 26, 2007
Early man 'couldn't stomach milk'
Working with scientists from Mainz University in Germany, the UCL team looked for the gene that produces the lactase enzyme in Neolithic skeletons dating between 5480BC and 5000BC. The paper will be Absence of the Lactase-Persistence associated allele in early Neolithic Europeans in PNAS. First LCT, then OCA2, and god knows what else? There is a limit in terms of what you can do with archeoDNA, but if the action went down in the last 10,000 years then it isn't such a sweat. One thing, the summary in Science is weird. Check it: The culture-historical hypothesis is that, shortly after the domestication of livestock, a few lucky farmers with a genetic anomaly hit white gold: nourishment via milk. Then, according to this theory, natural selection took over and these lactase-persistent folks proceeded to populate much of Europe with their milk-guzzling offspring. A competing hypothesis argues that ancient Europeans domesticated milk-bearing livestock because lactase persistence was already quite common in certain populations. Quite common? The area of the genome around LCT was hit by a hammer blow of selection less than 10,000 years ago, that's why researchers use it to check if their methods for detecting selection are working. The fact that independent events of lactase persistence exist among other populations via alternative genetic architectures seems to indicate clearly the power of gene-culture coevolution. What's up with Science doing a "look at both sides" framework when the evidence is so lopsided? I suppose selection could have been induced by a causative factor aside from milk, but I think we're verging into Humean skepticism at this point, denying the ability to discern causality at all.... Carl Zimmer has much more.... Related: Lactose tolerance by a different stroke. Lactose tolerance/intolerance. Are multiple lactose tolerance mutations surprising? Genes & culture & milk. Lactose tolerance, is it "dominant"? Milk digestion, it does a body good. Labels: Evolution, Genetics, Human Evolution
Sunday, December 10, 2006
Convergent adaptation of human lactase persistence in Africa and Europe:
A SNP in the gene encoding lactase (LCT) (C/T-13910) is associated with the ability to digest milk as adults (lactase persistence) in Europeans, but the genetic basis of lactase persistence in Africans was previously unknown. We conducted a genotype-phenotype association study in 470 Tanzanians, Kenyans and Sudanese and identified three SNPs (G/C-14010, T/G-13915 and C/G-13907) that are associated with lactase persistence and that have derived alleles that significantly enhance transcription from the LCT promoter in vitro. These SNPs originated on different haplotype backgrounds from the European C/T-13910 SNP and from each other. Nick Wade has an article in The New York Times. Labels: Human Evolution
Thursday, June 15, 2006
On the heels of the asinine review of Before the Dawn in Nature, I see Carl is linking to some recent papers that are coming out in regards to positive selection in our own storied lineage. I must say that the new one in Science is quite phat in its broad sweep. Pictures below the fold....
     Show me the money honey, you can sneer about incomplete evidence for selection on human lactase persistence, but saying it just ain't gonna make it so. Labels: Human Evolution
Readers know that I don't have a totally positive opinion of Before the Dawn, but this hatchet job in Nature is out of control, it stinks of "The Sociobiology Study Group." Full text below the fold, I'll let readers point out the issues....
In Your Own Image Labels: Human Evolution |
  Razib's Home Page GNXP Archives Interviews Blogroll Principles of Population Genetics Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism
Archives
July 2005 August 2005 September 2005 October 2005 November 2005 December 2005 January 2006 February 2006 March 2006 April 2006 May 2006 June 2006 July 2006 August 2006 September 2006 October 2006 November 2006 December 2006 January 2007 February 2007 March 2007 April 2007 May 2007 June 2007 July 2007 August 2007 September 2007 October 2007 November 2007 December 2007 January 2008 February 2008 March 2008 April 2008 May 2008 June 2008 July 2008 August 2008 September 2008 October 2008 November 2008 December 2008 January 2009 February 2009 March 2009 April 2009 May 2009 June 2009 July 2009 August 2009 September 2009 October 2009 November 2009 December 2009 January 2010 February 2010 Hello Movable Type archives August 11,2002 August 18,2002 August 25,2002 September 01,2002 September 15,2002 October 20,2002 December 08,2002 December 22,2002 December 29,2002 January 05,2003 January 12,2003 January 19,2003 January 26,2003 February 02,2003 February 09,2003 February 16,2003 February 23,2003 March 02,2003 March 09,2003 March 16,2003 March 23,2003 March 30,2003 April 06,2003 April 13,2003 April 20,2003 April 27,2003 May 04,2003 May 11,2003 May 18,2003 May 25,2003 June 01,2003 June 08,2003 June 15,2003 June 22,2003 June 29,2003 July 06,2003 July 13,2003 July 20,2003 July 27,2003 August 03,2003 August 10,2003 August 17,2003 August 24,2003 August 31,2003 September 07,2003 September 14,2003 September 21,2003 September 28,2003 October 05,2003 October 12,2003 October 19,2003 October 26,2003 November 02,2003 November 09,2003 November 16,2003 November 23,2003 November 30,2003 December 07,2003 December 14,2003 December 21,2003 December 28,2003 January 04,2004 January 11,2004 January 18,2004 January 25,2004 February 01,2004 February 08,2004 February 15,2004 February 22,2004 February 29,2004 March 07,2004 March 14,2004 March 21,2004 March 28,2004 April 04,2004 April 11,2004 April 18,2004 April 25,2004 May 02,2004 May 09,2004 May 16,2004 May 23,2004 May 30,2004 June 06,2004 June 13,2004 June 20,2004 June 27,2004 July 04,2004 July 11,2004 July 18,2004 July 25,2004 August 01,2004 August 08,2004 August 15,2004 August 22,2004 August 29,2004 September 05,2004 September 12,2004 September 19,2004 September 26,2004 October 03,2004 October 10,2004 October 17,2004 October 24,2004 October 31,2004 November 07,2004 November 14,2004 November 21,2004 November 28,2004 December 05,2004 December 12,2004 December 19,2004 December 26,2004 January 02,2005 January 09,2005 January 16,2005 January 23,2005 January 30,2005 February 06,2005 February 13,2005 February 20,2005 February 27,2005 March 06,2005 March 13,2005 March 20,2005 March 27,2005 April 03,2005 April 10,2005 April 17,2005 April 24,2005 May 01,2005 May 08,2005 May 15,2005 May 22,2005 May 29,2005 June 05,2005 June 12,2005 June 19,2005 June 26,2005 July 03,2005 July 17,2005 August 07,2005 Blogspot archives June 2002 July 2002 August 2002 September 2002 October 2002 November 2002 December 2002
10 questions for....
Parag Khanna James Flynn Jon Entine Gregory Clark György Buzsáki Heather Mac Donald Bruce Lahn A.W.F. Edwards Luigi Luca Cavalli-Sforza Joseph LeDoux Matthew Stewart Charles Murray James F. Crow Adam K. Webb Justin L. Barrett David Haig Judith Rich Harris Ken Miller Dan Sperber Warren Treadgold Armand M. Leroi John Derbyshire
Blogs
The GiveWell Blog Your Religion Is False Colby Cosh Steve Hsu Audacious Epigone Catallaxy Files Inductivist 2 Blowhards Genetic Future Agnostic Steve Sailer Dienekes Derek Lowe Razib Khan Razib at Comment is Free Secular Right Glenn Reynolds Jim Miller Kevin McGrew John Hawks Peter Fost Randall Parker Less Wrong Charles Murray Carl Zimmer EconLog Marginal Revolution
Principles of Population Genetics
Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism   Policies Terms of use © http://www.gnxp.com Razib's total feed: |
{kind=link}