
|
Thursday, February 26, 2009
Following up on this previous post on epigentics, I thought I'd point to a couple nice examples of using epigenetic information to obtain insight into basic biology.
The first is, I think it's fair to say, a landmark paper identifying a set of over a thousand likely functional non-coding RNAs in mouse cell lines. The approach used here was epigenetic: the authors generated genome-wide maps of chromatin modifications known to mark promoters and transcribed regions, and screened out all the regions of the genome already known to be transcriptionally active. This left them with a set of putatively functional transcripts, which tended to be highly evolutionarily conserved (indicating function), and many of which they confirmed via other means to be novel long non-coding RNAs. The second is a nice paper demonstrating that one of the same epigentic marks used above to identify transcribed regions is present, in humans, mice, and nemotodes, preferentially on exons (rather than on the entirety of the transcribed region). As this mark is present only in genes that are being transcribed, the authors conclude that it is placed in conjunction with transcription, and likely in conjunction with splicing. They speculate about the role that this mark could play in gene regulation, but in general, this paper raises many more interesting questions than can currently be answered. Labels: Epigenetics, Gene Expression
Tuesday, February 24, 2009
Not news to many readers, I'm sure, but Nicholas Wade has a nice article on epigenetics and gene regulation. Some people in the article complain about the lack of a focused investment by the government in this area. I found this a little odd--isn't quite a bit of large-scale work being done by the ENCODE project?
Labels: Epigenetics, Genetics
Sunday, May 27, 2007
Redefinition / turnin' your play into a tragedy / exhibit level degree on the mic / passionately - Kweli
Nature has an Insight section up ostensibly about epigenetics starting with an article by Adrian Bird suggesting a re-definition. It's free. I think the term is useful to the extent that you can predict something about a phenomenon or mechanism by knowing that it is 'epigenetic'. Bird's suggestion is to make the term less useful and more inclusive. The last section is labeled 'Refining a definition' when in fact he is doing just the opposite: ...there might be a place for a view of epigenetics that keeps the sense of the prevailing usages but avoids the constraints imposed by stringently requiring heritability. The following could be a unifying definition of epigenetic events: the structural adaptation of chromosomal regions so as to register, signal or perpetuate altered activity states. This definition is inclusive of chromosomal marks, because transient modifications associated with both DNA repair or cell-cycle phases and stable changes maintained across multiple cell generations qualify. He wants to allow transience and yet use epigenetics to explain stable phenomena: A growing idea is that functional states of neurons, which can be stable for many years, involve epigenetic phenomena, but these states will not be transmitted to daughter cells because almost all neurons never divide. Without such epigenetic mechanisms, hard-won changes in genetic programming could be dissipated and lost;With this refinement, epigenetics is everything and nothing. The only thing you can infer about an epigenetic event is that it doesn't change the DNA sequence. Bird wants to claim that a unifying trait is that epigenetics is 'responsive' rather than 'proactive'. I don't understand. If you're going to introduce new terms, why not choose to bring into to broader use the distinction between meiotically and mitotically heritable or force people to be specific about which chromatin modification they are referring to instead of saying 'epigenetic modifications'? Responsive and proactive are more loosely defined concepts destined to muddy waters and lead away from insight. Labels: Epigenetics
Thursday, April 26, 2007
Trevor at Epigenetics News follows up on our recent discussions about the term's increasing popularity. He's cites several good reasons why we might expect to see the term more often, and one reason that may not be all that good:
With all of these high profile and highly funded areas becoming closely associated with epigenetics, is it any surprise that more researchers are finding ways to include their focus as part of "epigenetics"? If a large enough portion of researchers stretch the word to apply to their topic of interest eventually 'epigenetics' will just mean 'biology'. Labels: Again, Epigenetics
Sunday, April 15, 2007
It appears that I'm not the only one musing on the meaning of 'epigenetic'. This timely essay from Mark Ptashne in Current Biology, takes a hard line, even calling into question the connection between histone modifications and epigenetics.
As a glance at the literature will reveal, however, histone modifications - acetylation, phosphorylation, methylation, and so on - are now often explicitly called 'epigenetic modifications'. This despite the fact that, so far as I am aware, no histone modification has been shown to be heritable. This section might be relevant if you are looking to 'epigenetic mechanisms' for the stability and persistence necessary for a long-term memory mechanism: Why might one be pre-disposed to misuse the term epigenetic? The term is sometimes used in the context of "maintaining stable states of gene expression", as though some 'locked in' mechanism, involving histone and/or DNA modifications, were required for stable states of gene expression in eukaryotes. But, in the first instance, there would seem to be no such special requirement: lambda lysogens are essentially infinitely stable in the absence of the specific signal that inactivates the repressor, and this system - of course - involves no histones. Labels: Epigenetics
Tuesday, April 03, 2007
Speaking of epigenetics, here's a rather interesting phenomenon:
Environmental contamination by endocrine-disrupting chemicals (EDC) can have epigenetic effects (by DNA methylation) on the germ line and promote disease across subsequent generations. In natural populations, both sexes may encounter affected as well as unaffected individuals during the breeding season, and any diminution in attractiveness could compromise reproductive success. Here we examine mate preference in male and female rats whose progenitors had been treated with the antiandrogenic fungicide vinclozolin. This effect is sex-specific, and we demonstrate that females three generations removed from the exposure discriminate and prefer males who do not have a history of exposure, whereas similarly epigenetically imprinted males do not exhibit such a preference. The observations suggest that the consequences of EDCs are not just transgenerational but can be "transpopulational", because in many mammalian species, males are the dispersing sex. This result indicates that epigenetic transgenerational inheritance of EDC action represents an unappreciated force in sexual selection. Our observations provide direct experimental evidence for a role of epigenetics as a determinant factor in evolution. Labels: Epigenetics
Monday, April 02, 2007
Epigenetics. What is it? Let's ask some reputable sources.
From the glossary of a review in the current Nature Reviews Genetics: Epigenetic - Refers to mitotically or meiotically heritable changes in gene expression that do not involve a change in DNA sequence. From the Cell epigenetics review issue a month and a half ago: Epigenetics, in a broad sense, is a bridge between genotype and phenotype-a phenomenon that changes the final outcome of a locus or chromosome without changing the underlying DNA sequence. ... More specifically, epigenetics may be defined as the study of any potentially stable and, ideally, heritable change in gene expression or cellular phenotype that occurs without changes in Watson-Crick base-pairing of DNA. Both definitions suggest that, to be epigenetic, something must 1) be heritable and 2) not be attributable to DNA sequence. Imagine if you took that "ideally" very liberally and dropped the heritability requirement. Now a change in gene function must be attributable to something besides DNA sequence. At that point epigenetics becomes gene regulation. Living cells are constantly performing gene regulation in myriad ways. If we loosen the definition enough, epigenetic could mean signal transduction, regulation of protein synthesis and degradation, and any and all transcriptional modulation. Definitions can and do change. One thing I'd like to do in this post is generate discussion as to the most useful way to use the term epigenetics, so that we can communicate most efficiently about the various biological topics now gathering under its umbrella. Here is a third definition from David Sweatt and Jonathan Levenson: A third definition posits that epigenetics is the mechanism for the stable maintenance of gene expression that involves physically 'marking' DNA or its associated proteins. This allows genotypically identical cells (such as all cells in an individual human) to be phenotypically distinct (for example, a neuron is phenotypically distinct from a liver cell). The molecular and physical basis for this type of change in DNA or chromatin structure is the focus of this review. By this definition, the regulation of chromatin structure is equivalent to epigenetics. In this definition, they are equating a mechanism for epigenetics with epigenetics. Now we have the same term at two levels of analysis. If you're a Hofstadter fan you can probably think of a lot of mischief to stir up when you run into this situation, but that's not the real problem. The problem stems from the fact that we already know that epigenetics isn't restricted to the regulation of chromatin structure, so in simplifying the term to one set of mechanisms we are discarding information and excluding mechanisms that we may not have even discovered yet. A lot of the time, Sweatt and Levenson use the term 'epigenetic mechanisms' to mean regulation of histone modification and DNA methylation. If we mean chromatin structure, why not say that? Why do Sweatt and company trade in 'chromatin modification' for 'epigenetics'? It's not an entirely arbitrary thing to do. The best understood mechanisms for epigenetic phenomena are DNA methylation and histone modification, hands down, and if you search Pubmed for epigenetics you will have a very difficult time finding an article that doesn't involve some combination of these chromatin 'marks'. The most-studied instance of epigenetics is probably imprinting. It has something for everyone, from an exciting battle-of-the-sexes evolutionary explanation to a cellular mechanism veiled in mystery. Very briefly, imprinting occurs when, for instance, you get the exact same DNA sequence for Gene A from mom and dad (you get two of each gene in case you forgot), but only the Dad copy of Gene A is active. The Mom copy is imprinted and thus silenced. Some percentage of human genes are imprinted. I think it's less than ten percent, can't remember. How is the gene expressed differently if the DNA sequence is the same? Well, during egg-ogenesis Mom added a methyl group (CH3, not a very reactive group, just makes the molecule bulkier) to a bunch of the cytosines in and around Gene A, thus changing them to 5-methyl-cytosines. This is called DNA methylation. In this example the gene is maternally imprinted, but don't get it twisted. Dads can imprint too. DNA methylation can affect gene expression directly by hiding sequences that would normally be recognized by regulatory proteins or indirectly as there are proteins that recognize methylated DNA and can recruit chromatin modification complexes to that locus. By the way, if any of this is terribly interesting to you please go check out one or two of the billion reviews on this stuff as I am not an expert in this area and I'd hate to lead you astray. DNA is often packed up by wrapping it around proteins called histones like you wrap the string around your yo-yo before you put it in the drawer. Unwrapping DNA from the histones allows other proteins (like RNA polymerase) access to the DNA and generally means an upregulation of that gene's activity. You can attach or detach acetyl groups or methyl groups (or phosphate groups or ubiquitin groups or SUMO groups, histones must look like Katamari Damacy) to the histones to make them more or less attracted to the DNA. Histone acetylation (by HATs) and deacetylation (HDACs, histone deacetylases) are core mechanisms for switching a section of chromosome between transcriptionally active and inactive states. There is a complex relationship between DNA methylation and histone modifications. As a basic scheme, DNA methylation is more often associated with silent genes wrapped tightly around deacetylated histones. The two occur independently though. Many transcriptional activators play their part by dragging some histone modification machinery to the gene they are interested in, so histones are dynamically modified in many cases of transcriptional regulation, whereas DNA methylation has been thought to be a more stable, long-lasting modification. So, yes. Very often 'epigenetic mechanism' refers to DNA or histone modification, but there are other ways of passing information between generations of organisms or cells without touching the DNA sequence. Yeast can transmit information across generations in the form of prions, infectious protein conformations. Think of Ice-9 in Cat's Cradle. When prions touch other normally folded proteins of the same type, they can transfom the normal protein. So a protein with the exact same amino acid sequence can fold up two different ways, one of which (the prion form) can be passed on to the offspring of the prion-infected yeast. Thus, a phenotypic determinant is passed across generations with no help from the DNA sequence. Also, last year a French group reported an instance of epigenetics in which accumulation of some abnormal RNA molecules in mouse sperm is responsible for inheritance of a specific coat pattern. All that is to say is that there are lots of ways to transmit information across generations and cell divisions. I think the thing about chromatin modification that was initially appealing to Sweatt and colleagues in terms of memory was its apparent stability. Research into cell differentiation, the process by which your basic stem cell produces the diverse array of cell-types required, has implicated chromatin modification as a strong influence. The idea is that to make, for instance, a neuron, you have to permanently set up a certain transcriptional program to make things like neurotransmitter receptors, so you go ahead and permanently turn on those genes. In some sense you could extend cell differentiation to a highly refined level so that the cell types aren't broad classes like neuron and skin cell, but rather 'neurons storing memory A' and 'neurons storing memory B'. Following this train of thought or something like it, they began to look for signs of chromatin modification in response to learning. They discovered that after a hippocampus-dependent form of fear conditioning, histones in the hippocampus are modified. Acetylation and phosphorylation are high one hour after training, but return to baseline by 24 hours. Given that we know that many genes are upregulated in response to fear conditioning and that transcription factors that activate genes have a history of modifying histones it would have been quite a surprise if we hadn't seen any changes. At this point, I began to get a little unhappy with the papers coming out of the Sweatt lab because I felt like the histone-modification story wasn't telling us much that we didn't know. So far, the epigenetics in memory story has been presented in a theoretical framework emphasizing the stability of chromatin modifications and the persistence of long-term memories. Just so I don't mischaracterize, here is a quote from 2006: One question that has eluded neuroscientists is, How can the brain store information over the lifetime of an organism in the face of molecular turnover? This question is especially relevant for understanding a complex process such as cognition, which relies heavily on the ability to store and recall information for periods longer than the half-lives of most of the molecules utilized in these processes. Chromatin is the one structure that remains relatively constant in almost every cell of a metazoan. It is not surprising that many recent studies in the nervous system indicate that from invertebrates to mammals, chromatin is a dynamic structure that integrates potentially hundreds of signals from the cell surface and effects a coordinated and appropriate transcriptional response. More important, chromatin is perhaps the only structure in a neuron capable of such higher-level signal integration and information storage that is not continually turned over. To rephrase: Memories are persistent. Therefore the molecular mechanisms for memory are persistent. Most molecules don't last, so they can't do the job. Chromatin structure lasts, so it is a good candidate for long-term memory storage. Here are my various criticisms of this line of thinking: 1) You can't have your cake and eat it too. Is chromatin a dynamic structure changing in the face of new stimuli or is it a solid, stable basis for long-term information storage? Once a cell's epigenome has been modified to store a memory does it somehow get write-protected? It seems to me that there are some chromatin modifications that are highly stable (i.e. the ones that control cell differentiation) that should be very unresponsive to the day-to-day vagaries of cell signaling, otherwise you would have neurons de-differentiating all willy-nilly. Another separate set of chromatin modifications must respond to cellular stimuli and allow dynamic gene expression to fit the needs of the cell in the current situation. 2) A cell only has one epigenome, but encodes more than one memory. That's the wonderful thing about synapses; they allow multiple excitability configurations. An average neuron has thousands of synapses. Memory 1 can light up synapses X, Y, and Z and cause the cell to fire a burst of action potentials whereas Memory 2 lights up a different set of synapses causing the cell to fire at a slower, more consistent rate. Maybe when synapse X loses its ability to excite the cell you forget a little bit and the burst of action potentials is weaker. That's one scenario. The point is illustrated empirically in studies like Vazdarjanova and Guzowski, (2004), which shows that 35% of cells in the CA1 region of the hippocampus light up in response to a single experience, and there is some 15% overlap between cell populations responding to two different contexts. The point of that report is actually to show that representations are relatively non-overlapping, but they aren't so sparse that you can afford a cell per experience. Is there some way to propose chromatin modification as a long-term memory mechanism that doesn't make this sparsity requirement? 3) The molecular turnover problem was never a problem. Sure molecules turnover, so the memory can't be housed in a single individual molecule. Dendritic spines (structures forming excitatory synapses), on the other hand, can remain stable for as long as we can record them. All of the molecular constituents of the spine turn over, but the structure remains. All of your skin cells die and are replaced, but your body remains intact. It's not an issue. The only way that molecular turnover would be an issue is some critical mass of the molecules in a structure turned over at once. I can imagine some mechanisms involving the post-synaptic scaffolding and structurally induced biochemical states, but the mechanism doesn't matter for putting the molecular turnover issue to bed. We have observed neuronal structures remaining intact for long periods of time. Apparently there are parts of the cell that are stable besides the chromatin. I can maybe think of a way out of #2, but #3 seems very clear and very much in conflict with the theoretical framework for this work. So when the new Miller and Sweatt came out I was thinking, "This is just the histone papers again with DNA methylation." I skimmed it, saw some of the experiments I expected, and gave it a general thumbs down. Now that I've given it a few reads I think the paper has some relevant and really novel findings, a major one being that DNA methylation is regulatable in nondividing cells. The enzymes that methylate DNA are called DNA methyltransferases (DNMTs). They most commonly add the methyl group to the C in the sequence, CG. Remember that DNA is double-stranded so you get a CG - GC quartet of nucleotides at these preferential methylation sites. There are two major classes of DNMT in mammals: de novo and maintenance. Maintenance DNMTs make sure that if a portion of the genome was methylated in a dividing cell, say a neuronal precursor cell, then that portion will be methylated in the daughter cells produced by the division. During DNA replication just prior to cell division you have double stranded DNA containing one of the original strands (with methylated CGs) and one newly synthesized strand. A maintenance DNMT recognizes the dsDNA with only a methylated CG facing an unmethylated GC and makes them match. De novo DNMTs can make new methylation sites where there was no indication of methylation to begin with. Their major role is during the 'reset' of methylation states that occurs during gametogenesis and in the preimplantation embryo, and they have been considered relatively quiet in the fully developed adult. The first thing Miller and Sweatt did was to show that the level of de novo DNMT mRNA is increased after fear conditioning. They then injected DNMT inhibitors into the hippocampus after fear conditioning and blocked long-term memory consolidation. I initially thought this experiment was suspect because I didn't understand that DNA methylation is not expected to be part of a cell's normal function. I associated DNA methylation with histone deacetylation and my understanding is that histone acetylation states are very commonly altered to tweak this or that gene's level up or down. In reality, people have hardly tested the effects of DNMT inhibitors in non-dividing cells. The literature is full of attempts to selectively target cancer cells with DNMT inhibitors because they have a habit of dividing too much. You would still like a more specific manipulation, a way of inhibiting DNMTs at specific genomic loci would be nice, but the behavioral control experiments in this paper go a long way toward showing that the drugs are not simply killing off neurons. The inhibitors are only effective at a time point shortly after training, defining a maximum window of some 6 hours in which relevant genes are demethylated and remethylated or vice versa in response to behavioral training. They next showed regulation of methylation at genes that are positively and negatively associated with memory formation. Protein phosphatase 1, a 'molecular constraint on learning and memory', is transiently methylated leading to reduced levels, whereas reelin, which has been associated with synaptic plasticity shows reduced methylation levels and increased mRNA expression. This second finding is pretty important. The identity of the mammalian demethylating enzyme is unclear leading some to question if an active demethylase exists at all. Demethylation in dividing cells can be achieved by simply being lazy about methylating new DNA. According to Hermann et al. in 2004, the 'best evidence so far for active demethylation in vivo" was the rapidity of sperm chromatin demethylation after fertilization. I'd say reduced methylation at the reelin locus in non-dividing cells within an hour after behavioral training is quite strong evidence. In fact, this window of very rapid demethylation at a specific locus suggests a strategy for investigating the biochemistry of mammalian demethylation. It should be possible to trap any proteins that are associated with the reelin promoter in the hour or so after training using techniques like chromatin immunoprecipitation. Finally, they emphasize that the methylation changes are transient. Everything returns to baseline by 24 hours after conditioning. This very obviously contradicts the idea of DNA methylation as a memory storage tool. For some reason, the discussion section still won't let it go though. Look at this: This attribute is, however, consistent with the role of the hippocampus as a structure contributing to memory consolidation but not memory storage. It will be interesting to determine if the Crick/Holliday mechanism plays a role in perpetuating long-term changes in adult neurons in the cortex, at known sites of long-term memory storage. The hippocampus' role in various types of memory may or may not be temporary. This is a broadly debated issue and in my humble opinion, the folks making the case for a long-term role for the hippocampus are doing a fine job. Regardless, no one suggests that the hippocampus is only involved for less than a day. So there must at the very least be a mechanism besides DNA methylation that helps maintain the memory for a week or so. I spent a lot of time looking into ways to specifically affect DNA methylation at certain loci. I found a paper where they made chimeric DNMTs with a custom DNA-binding domain and inserted the recognition sequence for that domain into the area to be methylated. To achieve this in vivo with any sort of temporal specificity would require the combination of at least three transgenes either through traditional methods or injection of viral vectors. The temporal specificity (within 6 hours) would still not really be on point, so I guess this isn't the solution. So now we know that some genes are regulated by methylation state in response to learning. The obvious next question is which genes. How does the cell decide which genes to turn up and which ones to turn down? Is there a signature pattern of CGs that indicates memory repressor vs memory enhancer genes? Do transcription factors that are already implicated in memory-related gene expression recruit DNA demethylase activity? A broader question remains. Why should mRNAs get expressed in response to learning anyway? Why involve the nucleus when the action is out at the synapses? I'm convinced that its important, but for my money, local protein synthesis from pre-existing RNAs seems a more specific response to changes in input patterns. Does anyone reading have an explanation for the need to produce new RNAs so far from the locus of plasticity? Labels: Epigenetics, Memory
Sunday, March 25, 2007
I'm working on a follow-up to my knee-jerk negative reaction to Miller and Sweatt last week. Surveying the field, I found that Nature Reviews Genetics just dropped an issue focusing in epigenetics with a complementary website.
At least a couple of the reviews spotlight our recently acquired ability to assay DNA methylation states on a genome-wide scale. I'm reading the Jirtle and Skinner piece now. Go check it out and report back. Labels: Epigenetics
Friday, March 16, 2007
There's a new article by Miller and Sweatt in Neuron claiming that DNA methylation is a step in memory formation. They show methylation and demethylation of particular genes (reelin and PP1) following fear conditioning, and they show that inhibition of DNA methyl transferases (the name says it all) during the memory consolidation can disrupt memory. I hope now that they've started naming genes at which the methylation matters they'll study something more specific. The idea of inhibiting all DNA methylation in a cell for any length of time seems too blunt an instrument. The cells responsible for that memory could've just keeled over or radicaly changed because some cell cycle regulator got turned up too high for instance. They do show that animals can form a new, strong memory a couple days after administration of the drug, but in some cases, animals with a hippocampal lesion can perform these tasks. The nervous system can achieve learning (especially learning as important as conditioned fear) through many means, so seemingly normal behavior after an insult isn't that strong a control.
The scope of the processes disrupted by a DNMT inhibitor is indicated by this sentence from Miller and Sweatt: Many developmentally important processes utilize this "prima donna" of epigenetics (Scarano et al., 2005 and Santos et al., 2005), including gene imprinting, cell differentiation, X chromosome inactivation, and long-term transcriptional regulation (Bestor et al., 1988 and Okano et al., 1998). Sorry to be such a naysayer. It is an interesting hypothesis. It is likely the case that DNA methylation is regulated during memory formation. If we just think that memory will require transcriptional regulation then probably some DNA modifications will have to be done and undone. Cis-regulatory sequences that control a gene's level of activation can act through recruitment of histone acetyl transferases and other chromatin modifiers, so a precedent exists. But I don't think there is any special connection between epigenetics and memory. Memory requires cells to be cells and work properly, so it will require transcriptional regulation and DNA/chromatin modification. For any further connection, I think we have to start naming names. Labels: Epigenetics, Memory
Friday, February 23, 2007
The current issue of Cell is dedicated to the rapidly changing field of epigenetics, loosely defined at the ensemble of chromatin and DNA modifications that structure the genome and control gene expression. The only review I've worked my way through yet is this one: Timescales of Genetic and Epigenetic Inheritance.
According to classical evolutionary theory, phenotypic variation originates from random mutations that are independent of selective pressure. However, recent findings suggest that organisms have evolved mechanisms to influence the timing or genomic location of heritable variability. Hypervariable contingency loci and epigenetic switches increase the variability of specific phenotypes; error-prone DNA replicases produce bursts of variability in times of stress. Interestingly, these mechanisms seem to tune the variability of a given phenotype to match the variability of the acting selective pressure. Although these observations do not undermine Darwin's theory, they suggest that selection and variability are less independent than once thought.The authors limit themselves largely to bacteria and yeast, but they document a great deal of evidence that parameters like the mutation rate change in response to external stimuli, perhaps in almost a "directed" fashion. Some work in primates has shown a correlation between the amount of mutation raining down on a population and the amount of beneficial mutations that arise. Could "directed" mutagenesis be an explanation for this? The main points from the paper are summarized in the figure below; essentially, epigenetic inheritance is a way for a population to respond to environmental changes that occur on time scales shorter than those needed for genetic evolution. 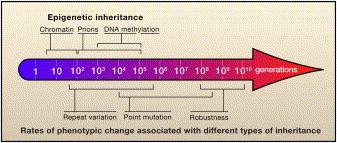 Labels: Epigenetics, Evolution |
  Razib's Home Page GNXP Archives Interviews Blogroll Principles of Population Genetics Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism
Archives
July 2005 August 2005 September 2005 October 2005 November 2005 December 2005 January 2006 February 2006 March 2006 April 2006 May 2006 June 2006 July 2006 August 2006 September 2006 October 2006 November 2006 December 2006 January 2007 February 2007 March 2007 April 2007 May 2007 June 2007 July 2007 August 2007 September 2007 October 2007 November 2007 December 2007 January 2008 February 2008 March 2008 April 2008 May 2008 June 2008 July 2008 August 2008 September 2008 October 2008 November 2008 December 2008 January 2009 February 2009 March 2009 April 2009 May 2009 June 2009 July 2009 August 2009 September 2009 October 2009 November 2009 December 2009 January 2010 February 2010 Hello Movable Type archives August 11,2002 August 18,2002 August 25,2002 September 01,2002 September 15,2002 October 20,2002 December 08,2002 December 22,2002 December 29,2002 January 05,2003 January 12,2003 January 19,2003 January 26,2003 February 02,2003 February 09,2003 February 16,2003 February 23,2003 March 02,2003 March 09,2003 March 16,2003 March 23,2003 March 30,2003 April 06,2003 April 13,2003 April 20,2003 April 27,2003 May 04,2003 May 11,2003 May 18,2003 May 25,2003 June 01,2003 June 08,2003 June 15,2003 June 22,2003 June 29,2003 July 06,2003 July 13,2003 July 20,2003 July 27,2003 August 03,2003 August 10,2003 August 17,2003 August 24,2003 August 31,2003 September 07,2003 September 14,2003 September 21,2003 September 28,2003 October 05,2003 October 12,2003 October 19,2003 October 26,2003 November 02,2003 November 09,2003 November 16,2003 November 23,2003 November 30,2003 December 07,2003 December 14,2003 December 21,2003 December 28,2003 January 04,2004 January 11,2004 January 18,2004 January 25,2004 February 01,2004 February 08,2004 February 15,2004 February 22,2004 February 29,2004 March 07,2004 March 14,2004 March 21,2004 March 28,2004 April 04,2004 April 11,2004 April 18,2004 April 25,2004 May 02,2004 May 09,2004 May 16,2004 May 23,2004 May 30,2004 June 06,2004 June 13,2004 June 20,2004 June 27,2004 July 04,2004 July 11,2004 July 18,2004 July 25,2004 August 01,2004 August 08,2004 August 15,2004 August 22,2004 August 29,2004 September 05,2004 September 12,2004 September 19,2004 September 26,2004 October 03,2004 October 10,2004 October 17,2004 October 24,2004 October 31,2004 November 07,2004 November 14,2004 November 21,2004 November 28,2004 December 05,2004 December 12,2004 December 19,2004 December 26,2004 January 02,2005 January 09,2005 January 16,2005 January 23,2005 January 30,2005 February 06,2005 February 13,2005 February 20,2005 February 27,2005 March 06,2005 March 13,2005 March 20,2005 March 27,2005 April 03,2005 April 10,2005 April 17,2005 April 24,2005 May 01,2005 May 08,2005 May 15,2005 May 22,2005 May 29,2005 June 05,2005 June 12,2005 June 19,2005 June 26,2005 July 03,2005 July 17,2005 August 07,2005 Blogspot archives June 2002 July 2002 August 2002 September 2002 October 2002 November 2002 December 2002
10 questions for....
Parag Khanna James Flynn Jon Entine Gregory Clark György Buzsáki Heather Mac Donald Bruce Lahn A.W.F. Edwards Luigi Luca Cavalli-Sforza Joseph LeDoux Matthew Stewart Charles Murray James F. Crow Adam K. Webb Justin L. Barrett David Haig Judith Rich Harris Ken Miller Dan Sperber Warren Treadgold Armand M. Leroi John Derbyshire
Blogs
The GiveWell Blog Your Religion Is False Colby Cosh Steve Hsu Audacious Epigone Catallaxy Files Inductivist 2 Blowhards Genetic Future Agnostic Steve Sailer Dienekes Derek Lowe Razib Khan Razib at Comment is Free Secular Right Glenn Reynolds Jim Miller Kevin McGrew John Hawks Peter Fost Randall Parker Less Wrong Charles Murray Carl Zimmer EconLog Marginal Revolution
Principles of Population Genetics
Genetics of Populations Molecular Evolution Quantitative Genetics Evolutionary Quantitative Genetics Evolutionary Genetics Evolution Molecular Markers, Natural History, and Evolution The Genetics of Human Populations Genetics and Analysis of Quantitative Traits Epistasis and Evolutionary Process Evolutionary Human Genetics Biometry Mathematical Models in Biology Speciation Evolutionary Genetics: Case Studies and Concepts Narrow Roads of Gene Land 1 Narrow Roads of Gene Land 2 Narrow Roads of Gene Land 3 Statistical Methods in Molecular Evolution The History and Geography of Human Genes Population Genetics and Microevolutionary Theory Population Genetics, Molecular Evolution, and the Neutral Theory Genetical Theory of Natural Selection Evolution and the Genetics of Populations Genetics and Origins of Species Tempo and Mode in Evolution Causes of Evolution Evolution The Great Human Diasporas Bones, Stones and Molecules Natural Selection and Social Theory Journey of Man Mapping Human History The Seven Daughters of Eve Evolution for Everyone Why Sex Matters Mother Nature Grooming, Gossip, and the Evolution of Language Genome R.A. Fisher, the Life of a Scientist Sewall Wright and Evolutionary Biology Origins of Theoretical Population Genetics A Reason for Everything The Ancestor's Tale Dragon Bone Hill Endless Forms Most Beautiful The Selfish Gene Adaptation and Natural Selection Nature via Nurture The Symbolic Species The Imitation Factor The Red Queen Out of Thin Air Mutants Evolutionary Dynamics The Origin of Species The Descent of Man Age of Abundance The Darwin Wars The Evolutionists The Creationists Of Moths and Men The Language Instinct How We Decide Predictably Irrational The Black Swan Fooled By Randomness Descartes' Baby Religion Explained In Gods We Trust Darwin's Cathedral A Theory of Religion The Meme Machine Synaptic Self The Mating Mind A Separate Creation The Number Sense The 10,000 Year Explosion The Math Gene Explaining Culture Origin and Evolution of Cultures Dawn of Human Culture The Origins of Virtue Prehistory of the Mind The Nurture Assumption The Moral Animal Born That Way No Two Alike Sociobiology Survival of the Prettiest The Blank Slate The g Factor The Origin Of The Mind Unto Others Defenders of the Truth The Cultural Origins of Human Cognition Before the Dawn Behavioral Genetics in the Postgenomic Era The Essential Difference Geography of Thought The Classical World The Fall of the Roman Empire The Fall of Rome History of Rome How Rome Fell The Making of a Christian Aristoracy The Rise of Western Christendom Keepers of the Keys of Heaven A History of the Byzantine State and Society Europe After Rome The Germanization of Early Medieval Christianity The Barbarian Conversion A History of Christianity God's War Infidels Fourth Crusade and the Sack of Constantinople The Sacred Chain Divided by the Faith Europe The Reformation Pursuit of Glory Albion's Seed 1848 Postwar From Plato to Nato China: A New History China in World History Genghis Khan and the Making of the Modern World Children of the Revolution When Baghdad Ruled the Muslim World The Great Arab Conquests After Tamerlane A History of Iran The Horse, the Wheel, and Language A World History Guns, Germs, and Steel The Human Web Plagues and Peoples 1491 A Concise Economic History of the World Power and Plenty A Splendid Exchange Contours of the World Economy 1-2030 AD Knowledge and the Wealth of Nations A Farewell to Alms The Ascent of Money The Great Divergence Clash of Extremes War and Peace and War Historical Dynamics The Age of Lincoln The Great Upheaval What Hath God Wrought Freedom Just Around the Corner Throes of Democracy Grand New Party A Beautiful Math When Genius Failed Catholicism and Freedom American Judaism   Policies Terms of use © http://www.gnxp.com Razib's total feed: |